 "
"
Team:Dundee/Project/PP1Capacities
From 2013.igem.org
| Line 17: | Line 17: | ||
<div class="span12"> | <div class="span12"> | ||
| - | <p>Microcystin’s toxic action lies in its ability to bind to and thus deactivate the human Protein Phosphatase-1 (PP1). Microcystin covalently binds to PP1 in a one-to-one relationship. Therefore, the larger the quantity of PP1 produced within a given chassis, the greater the | + | <p>Microcystin’s toxic action lies in its ability to bind to and thus deactivate the human Protein Phosphatase-1 (PP1). Microcystin covalently binds to PP1 in a one-to-one relationship. Therefore, the larger the quantity of PP1 produced within a given chassis, the greater the "mopping" potential .<br><br> |
The two options were explored for our bacterial mop: to export PP1 to the periplasm of <i>E. coli</i> and to export and then bind PP1 to the membrane surface of <i>B. subtilis</i>. To provide a crude estimate of ``maximum mop efficacy", we investigated the potential PP1-binding efficiency of both chassis option. | The two options were explored for our bacterial mop: to export PP1 to the periplasm of <i>E. coli</i> and to export and then bind PP1 to the membrane surface of <i>B. subtilis</i>. To provide a crude estimate of ``maximum mop efficacy", we investigated the potential PP1-binding efficiency of both chassis option. | ||
Revision as of 13:27, 23 October 2013

PP1 Packing
Microcystin’s toxic action lies in its ability to bind to and thus deactivate the human Protein Phosphatase-1 (PP1). Microcystin covalently binds to PP1 in a one-to-one relationship. Therefore, the larger the quantity of PP1 produced within a given chassis, the greater the "mopping" potential .
The two options were explored for our bacterial mop: to export PP1 to the periplasm of E. coli and to export and then bind PP1 to the membrane surface of B. subtilis. To provide a crude estimate of ``maximum mop efficacy", we investigated the potential PP1-binding efficiency of both chassis option.
Theory
When considering the maximum number of PP1 molecules physically compacted into the periplasm of E. coli or onto the membrane surface of B. subtilis, considerations for wasted space must be taken into account. This wasted space arises due to the impossibility of using all of the available space due to the physical configuration of PP1.
Before beginning such analyses, assumptions must be made regarding the shape and form of the PP1 molecules. To best describe the PP1 protein shape, without unnecessary over complication, PP1 was approximated as a sphere with a diameter equal to the width of the minimal osculatory cubic enclosure of the molecule. This approximation allows for the possibility of random orientations of the PP1 molecules and therefore gives an average result. The radius of PP1 is thus calculated as ~9.1nm.
In calculating the available volume of the E. coli or B. subtilis cell, the chassis were approximated as cylindrical bodies with hemispherical ends. Furthermore, the dimensions of E. coli and B. subtilis cells were taken as being approximately equivalent and were based upon the known dimensions of E. coli. “Each bacterium measures approximately 0.5 μm in width by 2 μm in length.”[2] .

The equations describing the volumes of spheres of radius r and cylinders of radius r and height h are given:
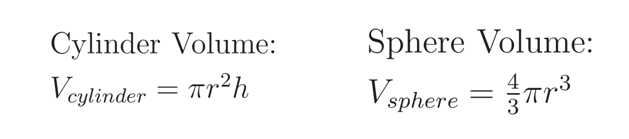
Making use of this approximation and these equations, the volume of an E. coli cell can be calculated as the volume of a cylinder of radius 0.25μm and height 1.5μm and the volume of a sphere of radius 0.25μm (the two hemispheres can be brought together and calculated as a single sphere).
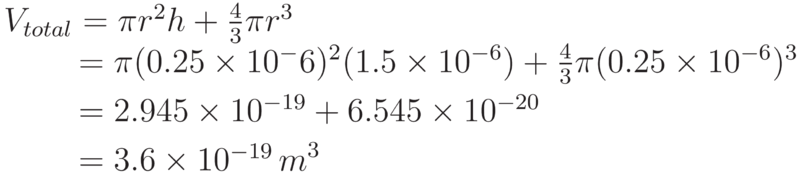
E. coli
For E. coli with a periplasmic width of 21nm, the periplasm volume can be calculated by considering the cell as two parts.
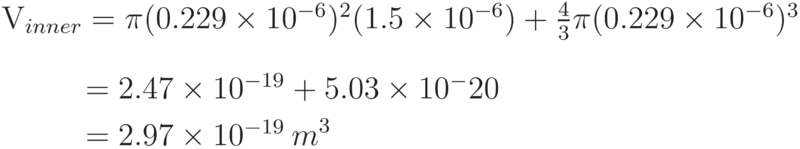
Therefore, the volume of the periplasm in E. coli is calculated to be:
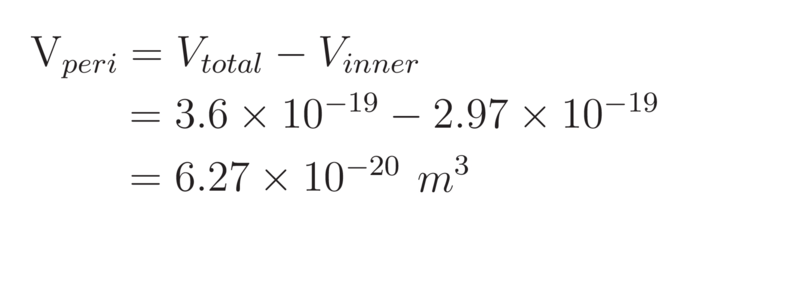
To determine the number of PP1 molecules that could be packed into the periplasm, we determined the volume of a single PP1 molecule. This volume can be taken as that of a sphere of diameter 9.1nm. However, when considering the volume that a single molecule will occupy, the space in between molecules must also be considered. To take account of this wasted space, a cubic approximation was made:
Thus, the volume of a PP1 molecule is assumed to be:
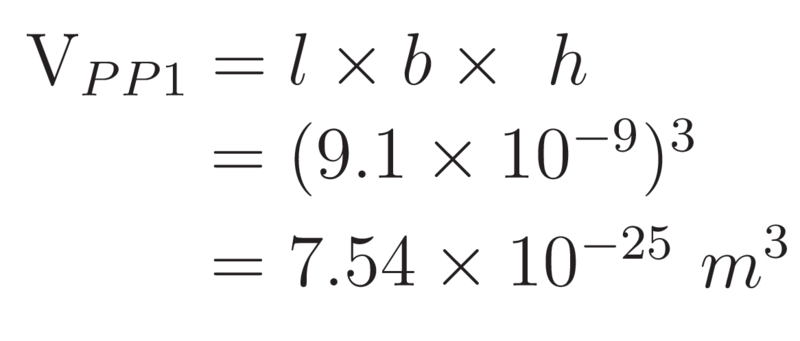
Hence, the maximum number of PP1 molecules that could be packed into the periplasm of E. coli can be approximated by:
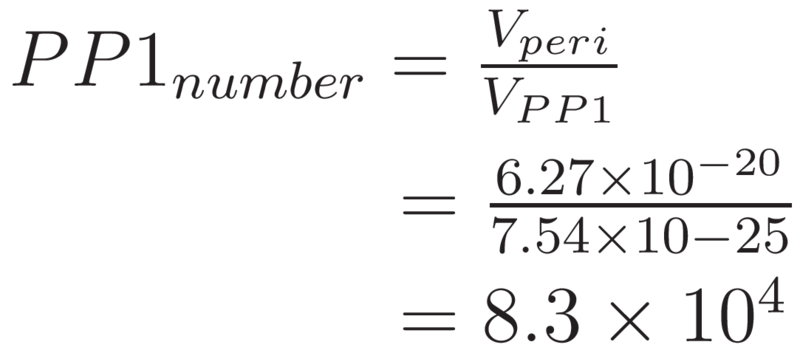
B. subtilis
When considering the packing of PP1 on the surface of B. subtilis, there is no defined volume for packing. B. subtilis is a Gram-positive cell and so the PP1 molecules are adhered in a monolayer to the surface of the cell membrane. Therefore, the approach to packing is different in this case.
In packing PP1 on the surface of the membrane, the number of PP1 molecules that can be placed “side-by-side” along the height of the membrane and around the circumference of the membrane was calculated.
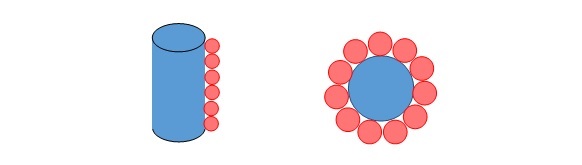
The number of PP1 molecules able to be stacked along the length of the cylindrical centrepiece of the cell (excluding the hemispheres) and around the circumference of the cylinder top:

Multiplying the above results gives the total number of PP1 molecules which can occupy the surface of the cylindrical midsection of the bacteria.
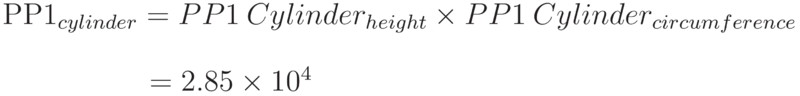
The number of PP1 molecules which could occupy the hemispherical ends can be calculated by approximating the hemispherical ends as flat circles. This approximation is appropriate due to the smaller contribution at these ends.
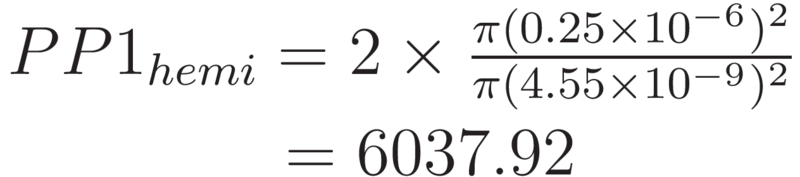
Thus, the total number of PP1 which could occupy the surface of B. subtilis can be calculated as the sum of the above.
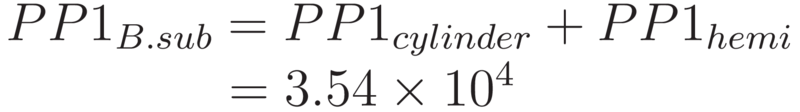
Conclusion
Having shown that E. coli has the potential to be the more efficient mop, this result was presented to the wet team in Week 1. The wet lab continued to explore the use of both species, but later in the project it was found that there were issues in exporting PP1 to the surface of B. subtilis. Hence our team focussed on E. coli as the chassis for the bacterial mop, in line with the original modelling prediction.
References
- [1] What is the hydrodynamic radius (RH)? Accessed: 18/09/2013:
http://ecoserver.imbb.forth.gr/pdf/hydrodynamic_radius.pdf - [2] Escherichia coli, PortEco. Accessed: 18/09/2013:
http://ecoliwiki.net/colipedia/index.php/Escherichia_coli - [3] Width of periplasm, BioNumbers. Accessed: 18/09/2013:
http://bionumbers.hms.harvard.edu/bionumber.aspx?&id=105387&ver=2 - [4] Protein serine/threonine phosphatase-1 (alpha isoform, type 1) complexed with microcystin-LR toxin, RCSB Protein Data Bank. Accessed: 18/09/2013: http://www.rcsb.org/pdb/explore/explore.do?structureId=1FJM#