 "
"
Team:SYSU-Software/model
From 2013.igem.org
Liuxiangyu (Talk | contribs) |
Liuxiangyu (Talk | contribs) |
||
| Line 14: | Line 14: | ||
<link href="https://2013.igem.org/Team:SYSU-Software/header.css?action=raw&ctype=text/css" rel="stylesheet" type="text/css" /> | <link href="https://2013.igem.org/Team:SYSU-Software/header.css?action=raw&ctype=text/css" rel="stylesheet" type="text/css" /> | ||
<link href="http://v2.bootcss.com/assets/css/bootstrap-responsive.css?action=raw&ctype=text/css" rel="stylesheet" type="text/css" /> | <link href="http://v2.bootcss.com/assets/css/bootstrap-responsive.css?action=raw&ctype=text/css" rel="stylesheet" type="text/css" /> | ||
| - | |||
| - | |||
| Line 166: | Line 164: | ||
<p> When α is the fold-change of activated transcription, n1a and n1r means the hill coefficient of the activator and repressor, respectively.</p> | <p> When α is the fold-change of activated transcription, n1a and n1r means the hill coefficient of the activator and repressor, respectively.</p> | ||
</section> | </section> | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
</div> | </div> | ||
</div> | </div> | ||
Revision as of 08:52, 27 September 2013

Overview
Mathematical modeling is the soul of our magical software "Computer Aided Synbio Tools", C.A.S.T. In our models, we take into account the intrinsic dynamics of different circuits or systems hierarchically by deterministic, stochastic and time delay models. Here are the highlights of our models and algorithms:
* We place great emphasis on various kinds of promoters, operons, coding parts, RBSs, terminators and apply different kinds of ODEs to describe their diverse behaviors in living cells;
* What our modeling tries to achieve is to creatively build a bridge between parts and modules in the Registry and widespread experimental data in the papers so that researchers can have a brand new perspective of how these parts are related to each other;
* Standardized output PoPS and RIPS is generated from raw experimental parameters, which represents our goal of establishing sets of standardized data;
* We create a new algorithm FoldChangeDecoder to calculate the fold-change in different cascades, widely used synthetic circuits, to make researchers have a more direct understanding of the regulatory networks;
*As the current focus of synthetic biology is readily combining modules into complex synthetic pathways[1], statistical and probability distribution models are also applied to estimate extrinsic variability among synthetic systems.
Intrinsic dynamics
Parameters

Deterministic models
The model mainly emphasizes on leakage rate of different promoters, the mRNA degradation term, and the strength of transcription, or called the rate of production. Also, transcription factors, such as repressors and activators, and coregulators are introduced to describe the dynamics of operons. In addition, the target DNA sequence is placed on Copynumber (CN) plasmids and some parameters are defined, for example, TS means the transcription strength of promoters while DeRNA represent the degradation rate of mRNA. Besides, we apply different levels of Hill input functions to model the behaviors of these regulators.
1. Constitutive Promoter
Transcription:
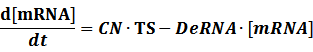
Translation:
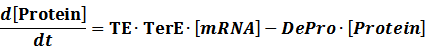
(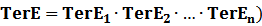
According to the iGEM Registry, the standardized input and output PoPS and RIPS can be given by the equations below based on our models:
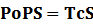
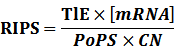
At steady level, the concentration of mRNA and Protein do not change.
Therefore, we have:
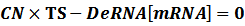
Thus,

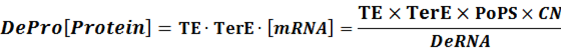
When there is a standardized output, we have:
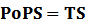
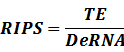
2. Inducible Promoter
2.1 one operon, positive or negative inducible
The first level: no transcription factor is added.
Here, we would introduce the leakage rate in order to estimate the production rate of the inducible promoters when no repressor or activator binds to the operon. Please note that, in negative inducible promoter, the leakage rate (LR) is almost as same as the transcription strength(TS) of the promoter in this situation.
Negative:
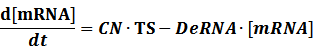
Positive:
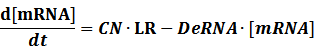
The second level : Promoter+n1 repressors or activators
(1) Negative inducible promoter
In repression of a promoter, n repressor proteins, and nR, bind together to the operator, a DNA site of the promoter, and thus there is a decrease in the rate of transcription of the promoter, so we have the basic kinetic equations
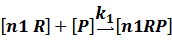
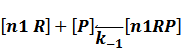
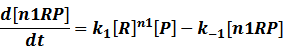
In steady state,
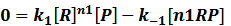
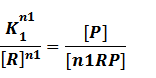
where
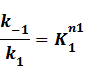
Suppose that the whole promoter regions,[PT], consist of active regions and inactive regions while there are two repressors, both active and inactive, can blind with regions, transcription can be only occurred in active regions.
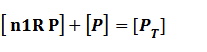
Thus, we could conclude that the promoter activity can be defined as (exclude the leakage part)
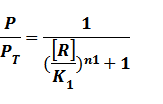
And we can find the ODE
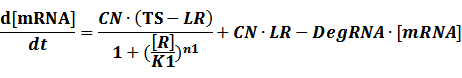
(2) Positive inducible promoter>
When the specific region is bound to n1 activators,
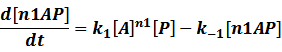
Similarly, in steady state,
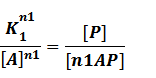
In our model, the difference between positive and negative regulation is that positive regulated circuits is for the regions that blind with active activators and can perform transcription.
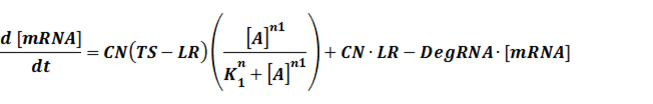
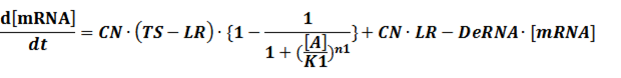
The third level: Promoter+n1 repressors or activators+n2 inducers or corepressors.
There are four cases at this level:
(1) Repressors + Inducers: every repressor is inactivated by n inducers;
Similarly,
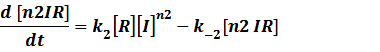
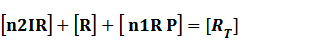
Because [n1RP] << [n2IR]+[R], we can apply
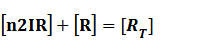
In steady state,
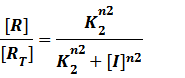
And
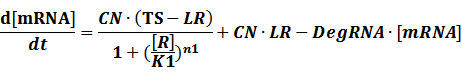
At last we can deduce that
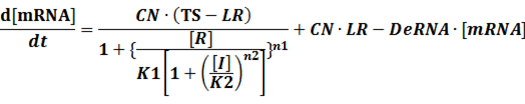
(2) Repressors + Corepressors: every repressor is activated by n corepressors;
Similarly,
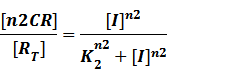
We assume that [n2CR] takes part in the repression of the promoter. Therefore, we have
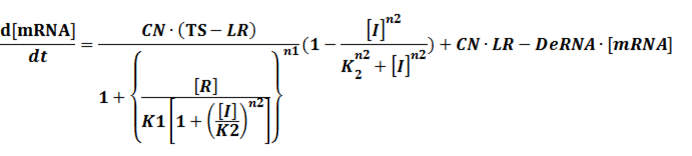
To simplify it,
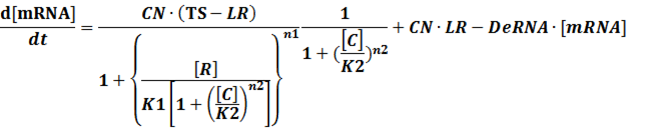
(3) Activators + Corepressors: every activator is inactivated by n corepressors;
Like (1), we can deduce that
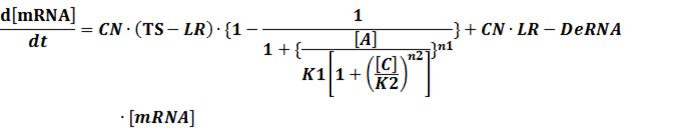
(4) Activators + Inducers: every activator is activated by n inducers;
We assume that [n2IR] also takes part in the activation of the promoter, so
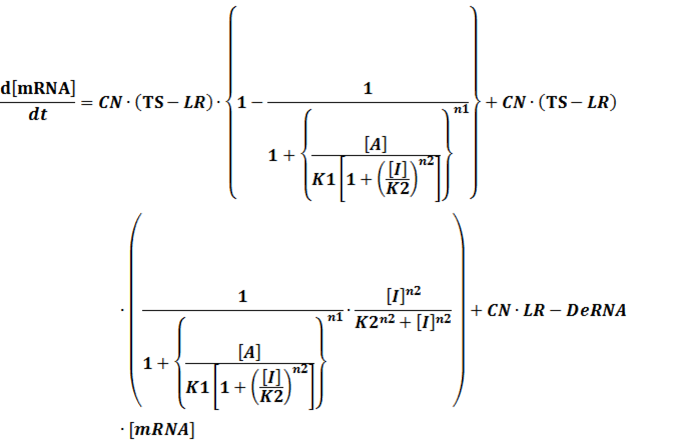
To simplify it,
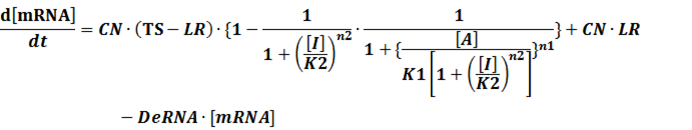
In conclusion, the output PoPS in these four cases can be defined as:
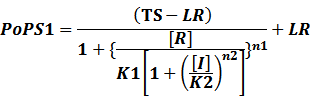
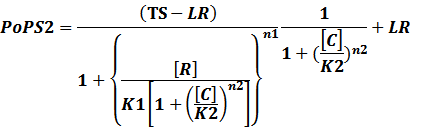
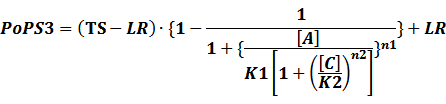
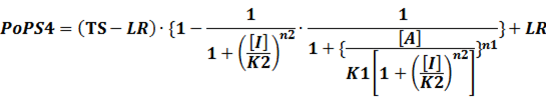
Note that when there is no inducer or corepressor, [C] and [I] would be 0. However, the standardized output RIPS always remains the same. Thus, we have
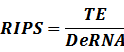
2.2 two operons O1 and O2 (double promoter)
It is common for prokaryotic cells to have two operons in a specific DNA site and there are also many useful double promoters in the iGEM Registry database. For example, BBa_I739104 is a double promoter (LuxR/HSL, positive / P22 cII, negative). In order to model these circuits, several rules and situations have to be considered:
(1)If both O1 and O2 host repressors, they can bind either cooperatively or not. Cooperativity between repressors implies an increase of the repressor affinity towards O2 when O1 is occupied;
(2)If both O1 and O2 host activators, they can bind either cooperatively or give rise to synergistic activation, which increases the promoter strength with a consequent increment of the transcription initiation frequency;
(3)O1 hosts an activator, while O2 hosts a repressor, which has proved to be easier for oscillator-building;
(4)Just to be simple, we do not consider the complete competition and cooperativity among the inducers or corepressors.
Like the example BBa_I739104 mentioned above, we can deduce the following equation for the transcription process:
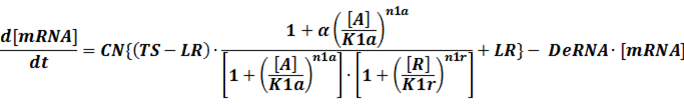
When α is the fold-change of activated transcription, n1a and n1r means the hill coefficient of the activator and repressor, respectively.
