"
"
Team:UCSF/Modeling2
From 2013.igem.org
| Line 34: | Line 34: | ||
| - | <div id="photos | + | <div id="photos"> |
| - | <img style="height:400px;margin-top: | + | <center><img style="width:950px;height:400px;margin-top:40px;margin-left:0px" |
| - | src="https://static.igem.org/mediawiki/2013/5/5f/ConjModel_UCSF.png"> </div> | + | src="https://static.igem.org/mediawiki/2013/5/5f/ConjModel_UCSF.png"> </center></div> |
| Line 53: | Line 53: | ||
</div> | </div> | ||
| - | |||
| - | |||
| - | |||
| - | |||
<div id="photos"> | <div id="photos"> | ||
Revision as of 01:45, 29 October 2013


The primary goal of the modeling portion for the CRISPRi conjugation project is to create a model that will help us identify the behavior of our transferable targeting system and identify critical parameters, given our assumptions. We used this model to check whether our CRISPRi conjugation system would have the desired behavior under biologically relevant parameters.
This system is designed to be transferred from cell to cell, targeting a specific gene of interest within the community. We model the activity of three strains of E. coli, an engineered donor strain containing a conjugative plasmid with genes coding for dCas9 and a gRNA, a recipient strain containing the RFP gene (target gene) in its genome, and upon successful conjugation, a transconjugate strain that no longer expresses RFP as well as the community's total RFP concentration.
Our objective is to simulate this community over time to see if our engineered system causes RFP levels to decrease.
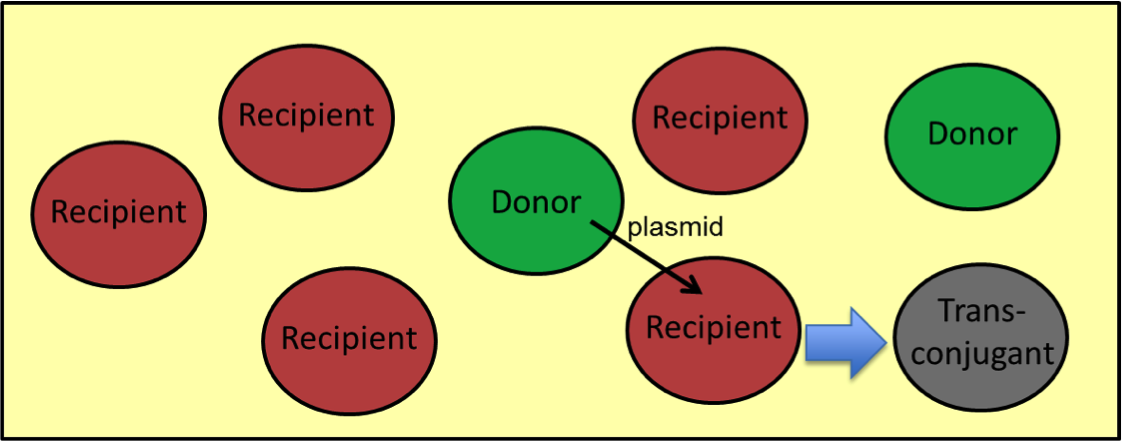

ASSUMPTIONS: While creating the model for our system, we made five assumptions in order to simplify some of the aspects of the model:
1) protein degradation is linear;
2) protein production is based on a hill function and also depends on inducer concentration;
3) repression is governed by a hill function and depends on the concentration of dCas9 and gRNA complex;
4) that the binding and unbinding of dCas9 and gRNA complex happens much faster than the production/degradation of gRNA and fluorescent proteins (the complex is at Quasi Steady State).
5) everything diffuses quickly throughout the cell so that our differential equations depends on the concentration at any given time.
EQUATIONS
Given these assumptions we have the following equations for the system:
Fluorescent proteins:
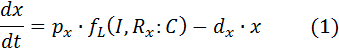
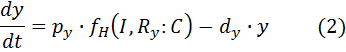
dx – rate of degradation of GFP based on the “low” function
dy – rate of degradation of RFP based on the “high” function
The production term consists of a function that relates inducer and repressor complex to mRNA production rate and conversion factor that relates mRNA production to protein production rate.
px – conversion factor (mRNA/protein) for x (GFP)
py – conversion factor (mRNA/protein) for y (RFP)
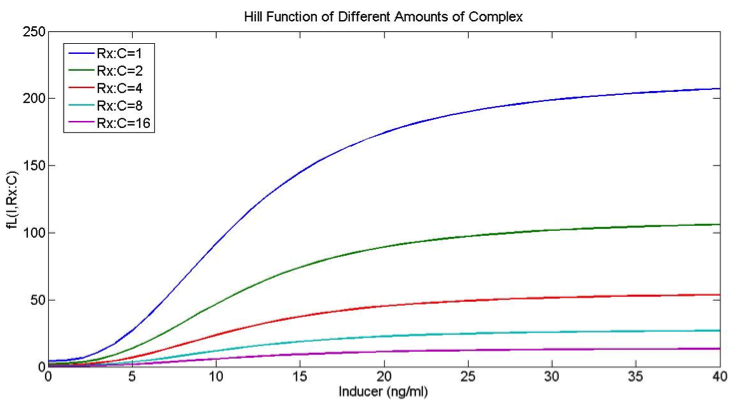
The mRNA production functions depend on the amount of inducer and the amount of repressor complex as follows:
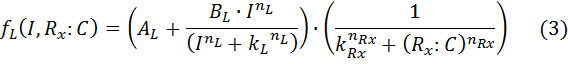
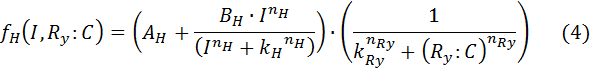
B – maximal expression level of promoter
k – Activation coefficient for low/high promoter
kR – half maximal effective concentration of R:C
n – hill coefficient for induction.
nR – hill coefficient for repression
I – Inducer concentration
R:C – repressor/dCas9 complex.
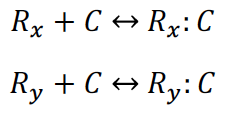
Given these chemical reactions, we can write the following equations for the gRNA/dCas9 Complex:
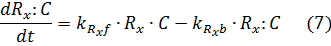
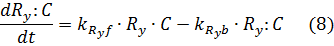
Under that assumption (setting equations (7) and (8) to zero – known as the quasi steady state assumption), we can solve for the complex in terms of the unbound repressor concentrations:
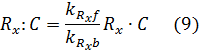
Where the amount of dCas9 available in the system is given by:
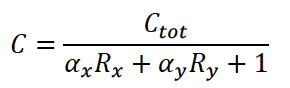
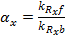
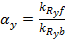
The equations for the gRNAs depend on the amount of the gRNAs that is produced, the degradation rate, and also the rate at which the gRNA complexes with dCas9. With the quasi-steady state assumption, the terms for complexing with dCAS9 drop out and the final equations for the gRNAs are similar to equations (1) and (2) for the fluorescent proteins:
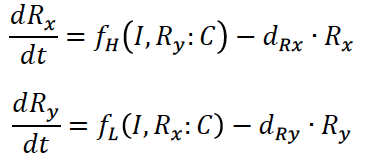
PARAMETERS
This model has many parameters, so in order for it to be more useful, we need to reduce the number of parameters that are undetermined. To accomplish this, we gathered some values from literature and also did experiments to find other parameters (Table1).


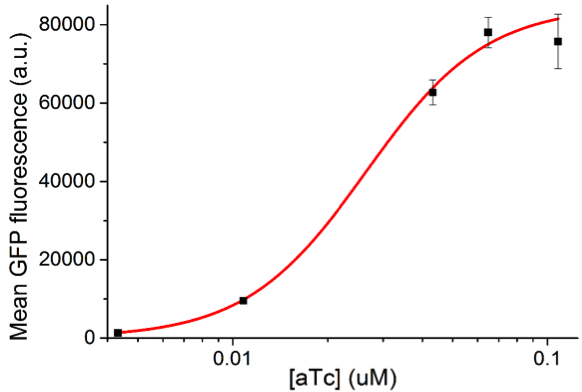
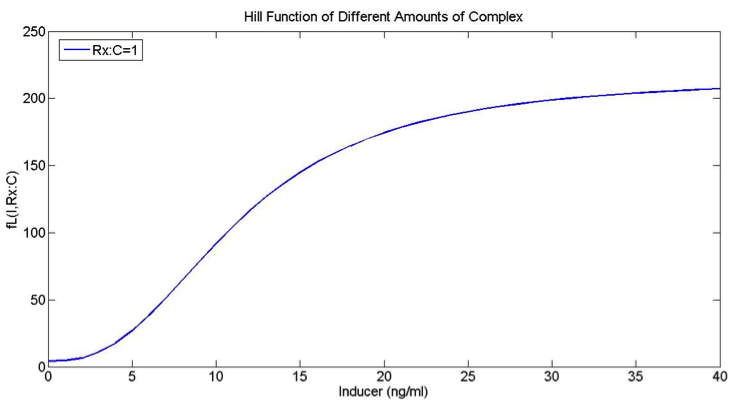
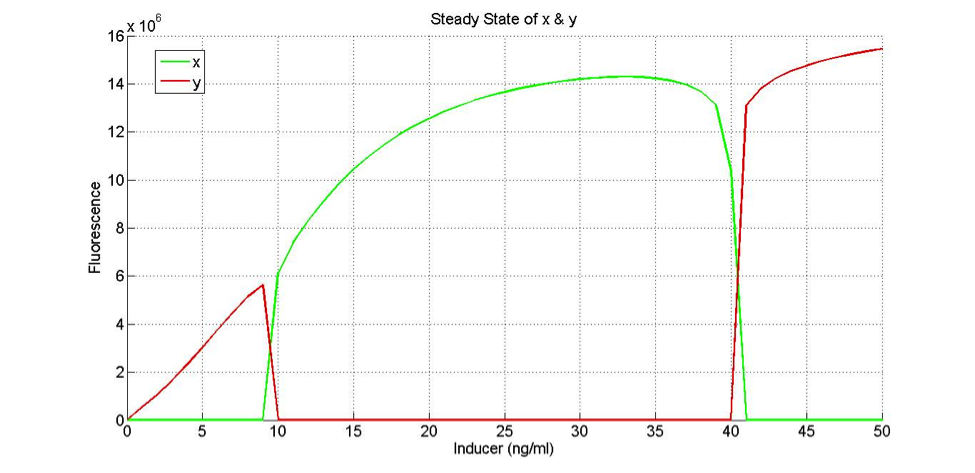
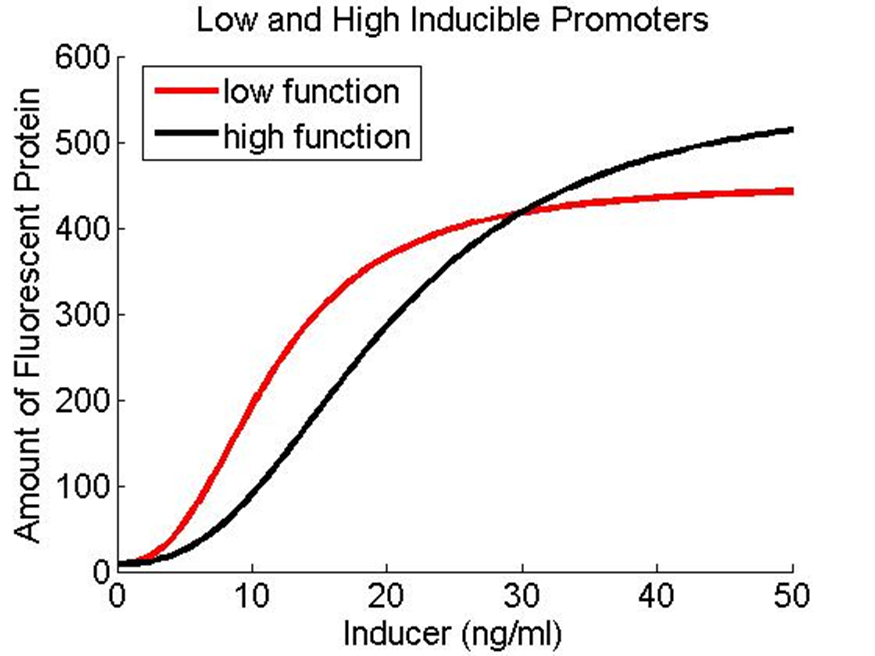
How does the model look with our actual “low” and “high” promoters?
If the only change in the low and high functions (FH and FL) is the K values (which determine the sensitivity of the promoters), then we don’t get our desired behavior. However, there are other parameters that might give us the desired behavior for the low and high promoters.
If we set BL to 443.7 and BH to 443.7*1.25, and if we set the half max values to kL = 11.45 and kH=17, the promoters have the following profile:
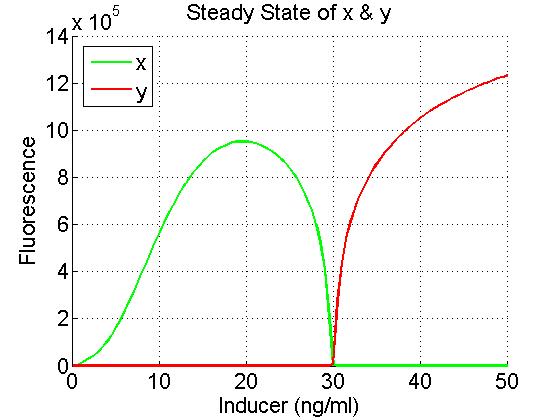
How does the system change when the hill coefficient is manipulated? In this first plot, the hill coefficients for both the low and the high function are the same number: 2.551. This number is the one we determined from our experimental data.
nL = 2.551
nH = 2.551
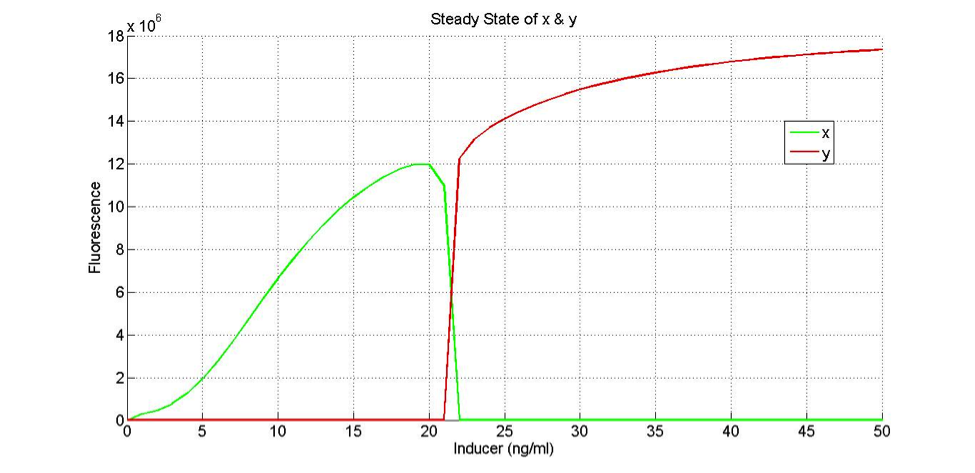
nL = 2.551
nH = 1.551