 "
"
Team:Valencia Biocampus/Project
From 2013.igem.org

Project Overview
Why E.coli
Escherichia coli is a model organism widely used in fields as microbiology, molecular biology and genetics. A result of that is the great range of genetic manipulation techniques that can be found related to this Gram negative bacterium. There is also a lot of information about the biochemistry and genetics of this microorganism. All this knowledge allows to work easily with it in the lab and to perform assays testing different conditions that will be tried out in other organisms unstudied in such depth. Moreover, E.coli is the main food of C.elegans, organism that is able to incorporate nucleic acids from the bacteria, RNA, to its cells (Mello et al., 2004). This is the usual mechanism of transformation of the worm, introducing exogenous genetic material from the bacteria altering the expression of the worm’s genome in order to make modifications of interest.
Controlling the mechanism
The role of E.coli in our project is to carry out the synthesis of an iRNA for inducing the social feeding behavior of Caenorhabditis elegans , the clumping. To achieve this, we cloned the biobrick Bba_K1112000 in E. coli, XL1-Blue strain (fig. 1).

Fig. 1.
- pSB1C3 is a high copy number plasmid (RFC [10]) carrying chloramphenicol resistance.
- The replication origin is a pUC19-derived pMB1 (copy number of 100-300 per cell).
- pSB1C3 has terminators bracketing its MCS which are designed to prevent transcription from *inside* the MCS from reading out into the vector. The efficiency of these terminators is known to be < 100%. Ideally we would construct a future set of terminators for bracketing a MCS that were 100% efficient in terminating both into and out of the MCS region.
This construction consists of the E.coli fadA promoter, which is activated in the presence of fatty acids (Clark, 1981), and the antisense sequence of the mRNA FLP-21 from C.elegans which encodes a protein involved in the solitary feeding behaviour of the worm, the formation of the dsRNA complex inhibits the expression of this protein, inducing the social feeding behavior (fig. 2).

Pseudomonas putida and PHA production
Pseudomonas putida is a gram-negative bacterium that is found in most soil and water habitats where there is oxygen. Its diverse metabolism and its capacity to break down organic harmful solvents (it has many genes involved in degrading aromatic or aliphatic hydrocarbons) in contaminated soils make this microorganism irreplaceable for research studies in the field of bioremediation but also for biosynthesis of value-added products. In addition, Pseudomonas putida has several strains including KT2440, the one we have worked with.
For example, it is capable of converting styrene oil into the biodegradable plastic PHA. This helps to degrade the polystyrene foam which
was thought to be non-biodegradable. Styrene is a major environment toxic pollutant released from industrial sites. The conversion to PHA allows the cure
of styrene pollution but it is also beneficial to society because of its applications in tissue engineering.
PHA is also environmental friendly and has a long self-life therefore it is also used in everyday items. Unlike styrene, PHA can break down in soil or water.
Within Pseudomonas putida, PHA accumulates under unbalanced growth conditions as a means of intracellular storage, storing the excess of carbon and energy. These PHA polymers are synthesized by enzyme PHA synthase which is bound to the surface of the PHA granules and uses coenzyme A thioesters of hydroxyalkanoic acids as substrates.

The role of P. putida in the Synthetic symbiosis that we have designed is to be carried by C. elegans to hotspots of interest where would produce bioplastic PHA.
An overview about our nematode
One of the main characters of our work is a nematode known as Caenorhabditis elegans, from the Rhabditidae family. It was first used as an experimental model in Developmental Genetics studies and nowadays is also used in other fields such as Clinical Biology, Neurobiology and Cell Biology, being a good model to study Alzheimer disease, obesity, diabetes and aging, among others.
Another interesting thing is that it feeds on Escherichia coli. Its “favorite” strain is OP50, although we checked that it’s also able to feed on XL1-Blue strain, the one that we used in all our molecular biology experiments.
Some advantages of C. elegans when compared with other model organisms are:
- Lifespan ranges between 2 and 3 weeks, so experimentation times are reduced.
- Its maintenance and study is cheap and simple (it is transparent, which facilitates microscopic observation).
- It is very small (1 mm), so it is possible to carry out experiments with a huge number of worms in a small Petri dish having a great statistical support.
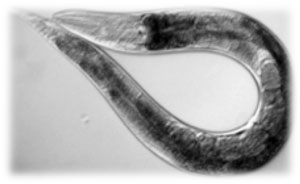
Why use C. elegans as a transport?
When we were considering on creating a new system for the transport of bacteria, we found different key advantages that made the nematode the best option. For example, C. elegans is able to move very fast around solid substrates (soil is, actually, its natural habitat) and also in agar, so it’s very useful for both lab experiments and real-environment tests.
Its movement, in addition to being very fast, has two modes: random and directed.
When there is no attractant in the medium, C. elegans moves doing uncoordinated movements in several directions in what is known as 'random walk'.
The situation changes when there is an attractant in the medium. Here, our 'transport' begins to direct his movements to the focus of the substance (volatile or soluble) which acts as an attractant in a process known as 'chemotaxis' thanks to the amazing smell of our nematode. This allows us to ‘guide’ the nematodes towards defined spots in irregular substrates.
Chemotaxis is the foundation to guide the transport of bacteria and is therefore the focus of experimentation with C. elegans, with the aim of finding the best attractant. This ability makes C. elegans a perfect 'bus' for bacteria. (simuelegans online here)
Moreover, we found two pathogens (Yersinia pestis and Xhenorhabus nematophila) with the ability to form biofilms on C. elegans thanks to the proteins of the operon hmsHFRS. When genetically-engineered strains of commonly used bacteria such as Escherichia coli or Pseudomonas putida express this hmsHFRS operon, they have the 'ticket' to travel: they are able to adhere to the worm’s surface by means of forming a synthetic biofilm.
Expanding our knowledge about C. elegans...
Once we knew that C. elegans was the best option, we began to discover interesting things for further study.
There are several strains of our nematode. The one that drew our attention was called 'N2', which had a fairly interesting behavior: under normal conditions, it eats individually, whereas under certain conditions (such as starvation), a social feeding behavior known as “clumping” is induced. But this phenomenon can also be induced if the expression of some particular genes is interfered. This fact gave us the opportunity to develop the first artificial symbiosis between worms and bacteria, based on the manipulation of the behaviour of C. elegans by simply nourishing it with transformed E.coli able to synthesize the iRNA.
With it, while C. elegans acts as transport, bacteria return the favour giving it the ability to eat in company.

Results
 The Riding
The Riding
The Riding
The first objective of our project was to genetically engineer bacteria so they can be able to “ride” the worms. Bacteria are not able to move fast in through solid or semi-solid substrates but they are very interesting from a biotechnological point of view, that is the reason why we though in this innovative mean of transport.
To achieve our goal, we constructed a BioBrick (see part: BBa_K1112001) consisting in the coding sequence of the hmsHFRS operon, an adhesion operon natural from Xenorhadbus nematophila which allows the formation of a biofilm on the nematode S. carpocapsae, under the control of a nitrogen sensitive promoter (pGlnA, characterized by the 2012 Valencia Biocampus iGEM team).
Our original idea was to transform Pseudomonas putida with the BioBrick, but the efficiency of the transformation was too low, so we decided to try with E. coli. Pseudomonas transformation with the operon could not be directly achieved with the vector in which the BioBrick was synthesized (pUC57 vector) so that we tried to insert the BioBrick in a vector with origin of replication in both Pseudomonas and E.coli, the pIZ1016 destiny vector (6,5Kb).
The mechanism of the construction works as follows: with low nitrogen in the media (0,6 g/L), the promoter is activated and hms genes are expressed, then the biofilm formation triggers; With high nitrogen concentrations (NH4(SO2)), as it is the case of nutrient-rich hotspots, the promoter is repressed and Pseudomonas should get out of the nematode. This process is what we call the riding.

Figure 1. Schematic representation of the BioBrick used for the riding. The effect of nitrogen concentration is shown in the drawings.
As you can see in the following electron micrography E.coli is able to colonize C.elegans’ cuticle under specific conditions:

 The Calling
The Calling
The Calling
Looking for an attractant
When we were considering setting up a transport of bacteria was thought to be necessary to have a 'destination', a place to go. That destiny would be a 'hot spot' on a heterogeneous substrate where Caenorhabditis elegans should lead right to that point the bacteria.Leveraging the powerful smell of the nematode, it was decided to try a number of attractants from various lists from web www.wormbook.org that could work as 'hot spot' of our experiment. Thus, using the C. elegans chemotaxis, we could direct transport.
The attractants experiment
The test would be carried creating our own plates on which half would be NGM unmodified and the other half part would be including soluble compounds before solidifying or after solidification in the case of volatiles. The list of modifications can be found in Fig. 1.

To place C. elegans on the plate, different cuts were made on a fresh plate of NGM, the resulting small pieces were placed in the exact center of the 50% -50% plates to determine which side of the nematode preferred, one per Petri plate.
The results after counting 2 replicates per attractant can be seen in Fig. 2.

From Table 2 it could rule out many of the attractants that were thought viable.
Volatile attractants were not a good choice to evaporate quickly (which is also limited to the field experiments).
Another impediment arose. Once had already performed the experiments, it was decided that the promoter that will control the production of RNA interference in Escherichia coli would be controlled by nitrogen, amino acids had to be discard as attractants; it would modify controlled expression of E.coli.
That left the MgSO4 and hypoosmotic media as potential attractants.
Attractant final choice
Because the hyposmotic medium could interfere with the proper growth of bacteria (food of our nematode), the final decision was to choose the MgSO4 as attractant.The question that arose at that time was: How we can multiply the amount of MgSO4 to increase efficiency?
MgSO4 efficiency
Once we had selected the most feasible attractant to the ‘transport’ for our experiments, we needed to know what could be the largest concentration of MgSO 4 in order to optimize the attraction of the nematode.To choose the concentration, were tested in a battery of increased concentrations regarding the initial medium (1 ml / L). Bearing in mind the results of the factor x2, we decided to test other factor concentrations: x3, x4, x5, x8 and x10; covering a range that does not exceed the concentration at which might affect the life of C. elegans or bacteria.
Moreover, approaching experiments with E. coli and Pseudomonas putida, these trials were testing the end media: half plate with NGM non-altered and half as PHA production medium for Pseudomonas and interference of E. coli.
Not knowing what fatty acid could activate transcription better, we tested two possibilities: oleic acid (named in the tables as PHAol) and octanoic acid (PHAoc). The concentration for each fatty acid was tested in E. coli choosing as better:
- 1.28 µl/ml of Octanoic acid.
- 2.58 µl/ml of Oleic acid.
You can see our results in figures 3, 4, 5 and 6. The two first counts were made after 3 hours and the next ones after 6 hours. We suppose at that time the worms can select their “favorite” half part gone across and their movement would be always in the same area. We prepare as control plates with the same composition but without MgSO4 in PHA media.

Final concentration choice
Once the test already performed and that the results seen by the factor of PHA and MgSO4 selected fatty acid give very scattered (there is probably repellent effect at high concentrations in the medium with oleic but reversed in octanoic) we make a selection of MgSO4 concentration for each medium.- If it is used oleic acid à Better results with 4ml/L MgSO4.
- If octanoic acid used à Best results to 10 ml/L of MgSO4.
The biggest problem in trying to have an effective attractant was how effective could be in presence of E. coli. It was therefore necessary to test the tradeoff between the value of 4ml / L MgSO4 and pair it with different concentrations of bacteria, high enough to feed the nematode but low enough to permit the attractive effect of MgSO4.
To find this point of commitment we prepare experiments in which the concentration of 4ml/L of MgSO4 is faced against different ODs from serial dilutions of a preculture of E. coli DH5a.
Table 7 shows the results. The count took place at 3h after the passing of fresh nematodes.
Best E. coli OD choice
An OD of 1 (minimum concentration of cells / volume) gives an attractive effect even better than expected (subsequent experiments try to see if there is synergy between E. coli and attractive factors MgSO4).To improve the approximation was decided to repeat the experiment with MgSO4 concentration and the chosen bacteria OD showing that the system works well. The results can be seen in figure 8.

 The Clumping
The Clumping
The Clumping
The base of this part is our engineered E.coli with a biobrick that is capable of changing the worm’s behavior.
Our C. elegans strain (N2 strain) has two feeding behaviors: Social and solitary feeding. Social feeding is known as clumping as you can see in this video:
Clumping is known to be induced under some conditions like temperature or starving, something that we had had considered during the entire project. However, it is also known how clumping is controlled from a genetic perspective and the main genes have already been described. We thought we could use this genetic approximation to control clumping under specific and controlled conditions. (view “Natural variation in a neuropeptide Y receptor homolog modifies social behavior and food response in C. elegans.” Bono M, Bargmann CI).
How do we induce clumping?
Our initial thought was to interfere the principal gene involved in this route: NPR-1. It is known that mutations in NPR1 convert a solitary strain into a social strain. Another option we finally chose was FLP-21, a gene that positively regulates NPR-1. We selected this one instead because it is not involved in so many vital processes. FLP-21 encodes a single FMRF amide-related neuropeptide that serves as a ligand forNpr-1, a G protein-coupled receptor that regulates social versus solitary feeding behavior in several Caenorhabditis species (see fig. 1).

So, this would be the final process: engineered E.coli that expresses FLP-21, whom RNA will interfere with the one codified by the worm because of the ingestion of bacteria. The formation of a complex of dsRNA will induce the elimination of Flp-21 transcripts by the RNA silencing pathway, which consequence is the induction of clumping (see figure 2). We also had to take into account that the induction of clumping by this mechanism is almost immediate, in comparison with natural clumping which takes longer depending on the external factors.

Why do we induce clumping for?
The main objective of the synthetic symbiosis that we designed was to detect hotspots of interest in irregular substrates and the induction of clumping was a good tool for it. With this natural mechanism, which we were going to control, worms kept in the desired places, then bacteria transported by C. elegans (Pseudomonas putida) were concentrated where their action was needed, in order to generate value-added products such as bioplastic PHA and the increase of worms in a specific location improved the image-based detection mechanism (go to Devices section)
Controlling the mechanism
The social feeding behavior needed to be controlled; it might be induced by conditions of interest as a tool for detecting hotspots in irregular substrates. We decided that fatty acids were going to be the inductors of the promoter (fadBp) that regulated the expression in the FLP-21 antisense sequence, in order to induce RNA interference (Fig.3). In the fatty acid rich media where this mechanism was activated, bacteria got off the worm, so this construction allows the induction of clumping in the spots where a biotechnological process will be carried out.

Choosing the right fatty acid
The E. coli’s biobrick promoter is induced by fatty acids, but those also induce the promoter of Pseudomonas putida in order to produce PHA. Some investigations showed that the induction of fadBp promoter by oleic acid is the most efficient (view “Regulation of fatty acid degradation in Escherichia coli: analysis by operon fusion” Clark D. et al) is higher when the fatty acid used is oleic acid but this is a long chained compound and that could affect the production of bioplastic (PHA). We decided to test the growth of E. coli and the production of bioplastic by P. putida using PHA media with oleic acid and octanoic acid in order to get the best results in both activities.Both species of microorganisms where grown in PHA production media with different concentrations of oleic and octanoic acids, maintaining a global concentration of fatty acids of 8mM, because it was the one where we found the highest growth of both E. coli and P. putida. Table 2 (below) summarizes the assays done.

The results of this assay showed that E. coli growth was better on PHA production media + 8mM oleic acid and the P. putida production of bioplastic was acceptable in this conditions. However, the highest production of bioplastic was on PHA production media+ 8mM octanoic acid one. So then, we made a Colonization assay and finally we determined that we were going to continue working using the PHA production media + 8mM oleic acid. Also, the ODs for plating both organisms were stablished as a consequence of this experiment. The “Choosing the right fatty acid” and the “Colonization” assays are explained at Building the bioplastic section.
Observing clumping induced by fatty acid rich media
We could see how clumping was done induced by the presence of 8mM of oleic acid (2.58 µl/ml ) which regulated the FadBp promoter of E. coli and as a consequence the expression the complementary sequence to the worms’ FLP-21 transcript, producing interference by RNA. In the plates there was a high concentration of E. coli from the serial centrifugations preculture 4ml XL1-Blue, 2 min at maximum rpm (2 times). The following images show the change of behavior of C. elegans, which at first were eating alone and then they were doing it in groups as it was planned (Fig. 4-6).

 The Building
The Building
The Building
We decided to include in the mechanism design a regulated production of a value-added product in the hotspots of interest were C. elegans would be attracted to. We got proffit of the natural capacity of Pseudomonas putida of producing bioplastic PHA (polyhydroxyalkanoates) by bacterial fermentation of sugar or lipids (fatty acids in this case), because we found that the directed production of this versatile material was interesting due to its easy detection (complexes of PHA with red Nile are fluorescent), its bright future in the field of biomaterials and because we were using this natural ability of bacteria as a tool.

To regulate the expression of the PHA cluster we used the glnA promoter that our team characterized the last year iGEM’s competition. (see https://2012.igem.org/Team:Valencia_Biocampus/Results1 ) for that, we optimized the concentration of ammonium sulphate for our Pseudomonas. We amplified the operon sequence and we sent it to the register (reference BBa_K1112002)
The role of the operon in our project
Our 2013 project is a proof of concept of the benefits that an artificial synthetic symbiosis between bacteria and nematodes can offer. The roles Pseudomonas play are two: firstly, they have been engineered to “ride” on C. elegans by the formation of a biofilm, but not stuffed with that, under the promoter glnA we stimulated the production of PHA, a value-added product that can be widely applied, for example:- For short disposable packaging items (personal hygiene products, surgical clothes).
- In upholstery.
- In the photographic and printing industry
- For textile industry since PHA can be processed into fibers.
- For biofuel production from PHA obtained from sewage sludge. This would combine two major advantages: the wastewater treatment and the generation of energy.
Choosing the right media
On the plates we were going to have both P. putida and E. coli and their promoters’ response towards fatty acids was different, so we had to make a media where E. coli could induce clumping and P. putida the production of bioplastic. In E. coli, the briobrick promoter for the expression of FLP-21 iRNA engineered in order to induce the social feeding behavior of C. elegans (Clumping) is induced by fatty acids, but not all the fatty acids are able to induce the same level expression (Fig. 2); oleic acid produces the highest activation of the promoter.

Whereas the standard medium for the production of PHA by Pseudomonas putida (composition reflected in Fig.3) employs octanoic acid (a short-chain fatty acid) as inducer. This fact made necessary to check if the modification of the fatty acid used, octanoic by oleic, would modify the bioplastic (PHA) production.

Otherwise, the medium for PHA production is tuned for the development of Pseudomonas putida, but not for the E.coli, so we had to test if this bacteria was able to grow in it. When we did this checking, we realized that the development of these bacteria did not occur in the PHA production media. However, with the addition of acetate (carbon source), the removal of the iron salt (added to the media to avoid the synthesis of a siderophore by Pseudomonas sp.) and, due to serendipity, with the double concentration of trace elementes, we got the media for the enteric bacteria growth.
Choosing the right fatty acid
Firstly, we tried to grow E. coli in the new PHA production media with different concentrations of octanoic acid. The same assays were done in parallel with P. putida in order to check which were the conditions where both microorganisms could grow and P. putida could produce enough quantity of bioplastic.After overnight incubation, the cultures were washed using sterile PBS and ODs600 were measured (Fig.4.). We also checked the PHA production by centrifuging the precultures, resuspending the pellet on PBS and adding Red Nile dissolved in DMSO, we could see the emission of fluorescence due to the interaction between the Nile red and the PHA through an UV transilluminator (Fig.5). By the results of the assays we chose to work with a 8mM concentration of fatty acids (Fig.4).

We also checked the PHA production by centrifuging the precultures, resuspending the pellet on PBS and adding Red Nile dissolved in DMSO, we could see the emission of fluorescence due to the interaction between the Nile red and the PHA through an UV transilluminator (Fig.5). By the results of the assays we chose to work with a 8mM concentration of fatty acids (Fig.4).

At this point we found that E. coli and P. putida could grow at 8mM of octanoic acid and that bioplastic (PHA) is also produced. However, because E. coli grows better in oleic acid and we wanted to check if P. putida could produce PHA if its phaC operon was induced by it, we grew both bacteria in different concentrations of octanoic and oleic acid but maintaining a constant concentration of fatty acids of 8mM. The assays done are explained in the table below (Fig.6).

We saw that E. coli had grown better with the increasing of oleic acid, getting at its maximum on experiment 9, even so, the quantity of pellet corresponding to 1 mL of preculture was scarce (and not visible at experiments 1 to 5). E. coli needed more time to grow in the PHA production media.
P. Putida had grown too, however we wanted to measure the production of bioplastic, we measured the ODs600 from the precultures of P. putida by centrifugating 2 mL of them and then washing them on 1 mL of PBS and resuspending the pellet in 500 μl of PBS. Then, dilutions on PBS were adjusted to the lowest OD600,corresponding to the experiment number 9 (8mM oleic acid). The growth of P. putida lower in PHA production media with a concentration of 8mM of oleic acid (Fig. 7-8).

The results showed that the growth of E. coli on 8mM of oleic acid was better than in any other condition, then experiment 9 conditions would be the best ones for the induction of clumping. However, even P. putida had produced bioplastic at all oleic acid concentrations, the highest production levels corresponded to experiment 1 conditions, which means in absence of oleic acid, just on 8mM of octanoic acid. Because of that, next assays were done following the conditions of experiment 1 and 9 (Fig.9).

After the optimization of the media, kinetics of the production of PHA and biomass generation were realized in relation to the time (Fig.10).

Colonization assays
For establishing the right media, the one in which E. coli could be able to grow and induce clumping and P. putida produce PHA and get to a balance between the two activities and for setting up some standard bases for doing an assay with the three microorganisms chosen for this synthetic symbiosis that our team proposes, we made some colonization assays. Different ODs600 of E. coli where plated in PHA production media in the conditions of both experiments 1 and 9 (Fig.6). There were two sets of plates:- 1) PHA production media + 8mM oleic acid
- 2) PHA production media + 8mM octanoic acid
In both of them (and their replicas) were plated the following E. coli ODs600 (Fig.11) and after let them grow for 48 hours, the plates were divided into 4 parts in order to plate different P. putida ODs600 (Fig.11). The huge difference between the ODs chosen for E. coli and P. putida was because we wanted to plate similar quantities of the Pseudomonas that C. elegans could afford to carry, taking into account that the worm size is 1mm, approximately.

Then the production of bioplastic was seen by the fluorescence of the Nile red joined to the PHA (the samples were prepared as in The right media). There were taken pictures of the plates but there was only growth of Pseudomonas in the first set of plates (PHA production media + 8mM oleic acid). There was a notorious growth and production of bioplastic in the plates where E. coli was plated with an OD of 0,8 an 1,0 and the production of bioplastic was higher at 0,03 and 0,05 OD600 of P. putida. (Fig.12).

Parts
These are the BioBricks we have designed, constructed, and characterized. We have submitted them to the Registry of Standard Biological Parts
| Works? | Name | Type | Description | Designer | Length | Structure |
|---|---|---|---|---|---|---|
| BBa_K1112000 | Regulatory | fadB promoter + FLP-21 iRNA | Pedro Luis Dorado Morales | |||
| BBa_K1112001 | Regulatory | pGlnA + hmsHFRS operon | Alba Iglesias Vilches | |||
| BBa_K1112002 | DNA | Cluster PHA | Alba Iglesias Vilches |
BBa_K1112000: fadB promoter + FLP-21 iRNA
This construction is made up by a fatty acid-sensitive promoter that directs the transcription of an iRNA responsible for the social or solitary behavior of C.elegans.
BBa_K1112001: pGlnA + hmsHFRS operon
We reused our nitrogen sensitive promoter from one of our last year constructs and we employ it to control the expression on the operon that triggers the formation of a biofilm over C.elegans.
BBa_K1112002: cluster PHA
This BioBrick contains the complete natural sequence that induces the production of PHA in Pseudomonas putida (KT2440).