 "
"
Team:HUST-China/Modelling/DDE Model
From 2013.igem.org
(Difference between revisions)
HUST LiChen (Talk | contribs) |
HUST LiChen (Talk | contribs) |
||
| Line 92: | Line 92: | ||
<img src="https://static.igem.org/mediawiki/2013/a/ab/Pathway_HUST.png"> | <img src="https://static.igem.org/mediawiki/2013/a/ab/Pathway_HUST.png"> | ||
<p class="small">Fig 1.The pathway of genetic oscillator</p><br /> | <p class="small">Fig 1.The pathway of genetic oscillator</p><br /> | ||
| - | The Arabinose Operon and the lac Operon is the core to the functioning of the oscillator. With the presence of Arabinose, dimeric AraC \cite{AraC_dimer}can induce the expression of downstream gene; On the other hand, with minor presence of IPTG, tetrameric LacI \cite{lac_tetramer}may suppress the expression of downstream gene. | + | The Arabinose Operon and the lac Operon is the core to the functioning of the oscillator. With the presence of Arabinose, dimeric AraC \cite{AraC_dimer}can induce the expression of downstream gene; On the other hand, with minor presence of IPTG, tetrameric LacI \cite{lac_tetramer}may suppress the expression of downstream gene. However, AraC and LacI cannot combined with promoters simultaneously(citation needed). Therefore, we have: |
$$D+2a\overset{k_1}{\underset{k_{-1}}\rightleftharpoons} D_1$$ | $$D+2a\overset{k_1}{\underset{k_{-1}}\rightleftharpoons} D_1$$ | ||
$$D+4r\overset{k_2}{\underset{k_{-2}}\rightleftharpoons} D_2$$ | $$D+4r\overset{k_2}{\underset{k_{-2}}\rightleftharpoons} D_2$$ | ||
where D stands for ratio of promoters which don't combine with any protein among all promoters, a for AraC protein(activator), r for LacI protein(repressor), $D_1$ for ratio of operons combined with AraC dimer, $D_2$ for ratio of operons combined with tetrameric LacI, $k_1$,$_{-1}$,$k_2$ and $k_{-2}$ are reaction rate constants. | where D stands for ratio of promoters which don't combine with any protein among all promoters, a for AraC protein(activator), r for LacI protein(repressor), $D_1$ for ratio of operons combined with AraC dimer, $D_2$ for ratio of operons combined with tetrameric LacI, $k_1$,$_{-1}$,$k_2$ and $k_{-2}$ are reaction rate constants. | ||
| - | + | We assumed that comparing with numbers of operons, that of protein is significantly large enough to ignore whose changes brought by combination of protein and operons. According to law of mass action, we had: | |
$$\frac{dD}{dt}=-k_1Da^2+k_{-1}D_1-k_2Dr^4+k_{-2}D_2$$ | $$\frac{dD}{dt}=-k_1Da^2+k_{-1}D_1-k_2Dr^4+k_{-2}D_2$$ | ||
$$\frac{dD_1}{dt}=k_1Da^2-k_{-1}D_1$$ | $$\frac{dD_1}{dt}=k_1Da^2-k_{-1}D_1$$ | ||
| Line 108: | Line 108: | ||
$$D_1 = \dfrac{a^2}{a_0(1+\frac{a^2}{a_0}+\frac{r^4}{r_0})}$$ | $$D_1 = \dfrac{a^2}{a_0(1+\frac{a^2}{a_0}+\frac{r^4}{r_0})}$$ | ||
$$D_2 = \dfrac{r^4}{r_0(1+\frac{a^2}{a_0}+\frac{r^4}{r_0})}$$ | $$D_2 = \dfrac{r^4}{r_0(1+\frac{a^2}{a_0}+\frac{r^4}{r_0})}$$ | ||
| - | What's worth mentioning is that both $a_0$ and $r_0$ are constant that are related to IPTG(mM) and Arabinose( | + | What's worth mentioning is that both $a_0$ and $r_0$ are constant that are related to IPTG(mM) and Arabinose(%). |
$$a_0 = \dfrac{(6.25+ara^2)(1+\frac{iptg^2}{3.24})}{100ara^2}$$ | $$a_0 = \dfrac{(6.25+ara^2)(1+\frac{iptg^2}{3.24})}{100ara^2}$$ | ||
$$r_0 = \dfrac{1}{2000000(\dfrac{0.19}{1+{\frac{iptg}{0.035}}^2+0.01})}$$ | $$r_0 = \dfrac{1}{2000000(\dfrac{0.19}{1+{\frac{iptg}{0.035}}^2+0.01})}$$ | ||
| Line 128: | Line 128: | ||
$$a\xrightarrow{\lambda f(x)}\varnothing$$ | $$a\xrightarrow{\lambda f(x)}\varnothing$$ | ||
$$r\xrightarrow{f(x)} \varnothing $$ | $$r\xrightarrow{f(x)} \varnothing $$ | ||
| - | where $d_{a/r}$, $f(x)$ and $\lambda f(x)$ are degradation rate constants. | + | where $d_{a/r}$, $f(x)$ and $\lambda f(x)$ are degradation rate constants. <br> |
According to law of mass reaction, we had: | According to law of mass reaction, we had: | ||
$$\frac{dR_a}{dt} = copy_a(k_3D_1+k_4D_2+k_5D)-d_{a/r}R_a$$ | $$\frac{dR_a}{dt} = copy_a(k_3D_1+k_4D_2+k_5D)-d_{a/r}R_a$$ | ||
| Line 136: | Line 136: | ||
$$\frac{da}{dt} = k_{fa}a_{uf} - \lambda f(x)a$$ | $$\frac{da}{dt} = k_{fa}a_{uf} - \lambda f(x)a$$ | ||
$$\frac{dr}{dt} = k_{fr}r_{uf} - f(x)r$$ | $$\frac{dr}{dt} = k_{fr}r_{uf} - f(x)r$$ | ||
| - | where $copy_a$ and $copy_r$ are plasmid copies that are transfected into E.coli. | + | where $copy_a$ and $copy_r$ are plasmid copies that are transfected into E.coli.<br> |
Transcriptional and translational processes of genes take time and consequently, protein that combined to promoters can be seen as those started transcription process before a specific time interval $\tau$. Thus we converted three ODEs into DDEs: | Transcriptional and translational processes of genes take time and consequently, protein that combined to promoters can be seen as those started transcription process before a specific time interval $\tau$. Thus we converted three ODEs into DDEs: | ||
$$D = \dfrac{1}{1+\frac{a(t-\tau)^2}{a_0}+\frac{r(t-\tau)^4}{r_0}}$$ | $$D = \dfrac{1}{1+\frac{a(t-\tau)^2}{a_0}+\frac{r(t-\tau)^4}{r_0}}$$ | ||
Revision as of 02:54, 23 October 2013








DDE MODEL
Methods
1. Establish DDEs based on mass action law;2. Investigate reasonable parameters set from previous researches;
3. Discuss DDEs' stability through examining their characteristic equations;
4. Determine the most effective factors in changing period;
Establishing DDEs
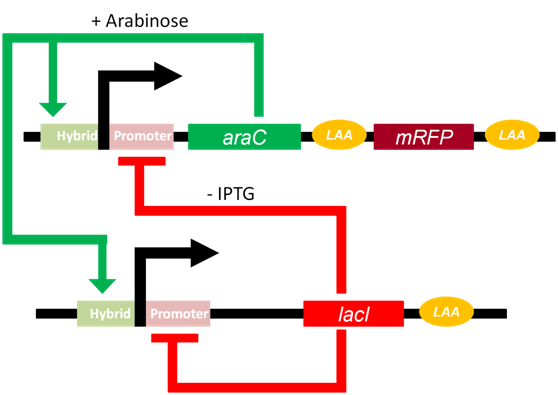
Fig 1.The pathway of genetic oscillator
The Arabinose Operon and the lac Operon is the core to the functioning of the oscillator. With the presence of Arabinose, dimeric AraC \cite{AraC_dimer}can induce the expression of downstream gene; On the other hand, with minor presence of IPTG, tetrameric LacI \cite{lac_tetramer}may suppress the expression of downstream gene. However, AraC and LacI cannot combined with promoters simultaneously(citation needed). Therefore, we have: $$D+2a\overset{k_1}{\underset{k_{-1}}\rightleftharpoons} D_1$$ $$D+4r\overset{k_2}{\underset{k_{-2}}\rightleftharpoons} D_2$$ where D stands for ratio of promoters which don't combine with any protein among all promoters, a for AraC protein(activator), r for LacI protein(repressor), $D_1$ for ratio of operons combined with AraC dimer, $D_2$ for ratio of operons combined with tetrameric LacI, $k_1$,$_{-1}$,$k_2$ and $k_{-2}$ are reaction rate constants. We assumed that comparing with numbers of operons, that of protein is significantly large enough to ignore whose changes brought by combination of protein and operons. According to law of mass action, we had: $$\frac{dD}{dt}=-k_1Da^2+k_{-1}D_1-k_2Dr^4+k_{-2}D_2$$ $$\frac{dD_1}{dt}=k_1Da^2-k_{-1}D_1$$ $$\frac{dD_2}{dt}=k_2Dr^4-k_{-2}D_2$$ With $a,a_0,r,r_0 > 0$, we found that eigenvalues $\lambda < 0$. Thus, when $t \rightarrow \infty$ we had$\frac{dD}{dt}=\frac{dD_1}{dt}=\frac{dD_2}{dt} = 0$. We also have $D+D_1+D_2 = 1$, so we arrive at: $$D=\dfrac{k_{-2}}{k_2r^4+k_{-2}+\frac{k_1k_{-2}a^2}{k_{-1}}}$$ $$D_1 = \frac{k_1a^2}{k_{-1}}D$$ $$D_2 = \frac{k_2r^4}{k_{-2}}D$$ Denote $a_0 = \frac{k_{-1}}{k_1}$ and $r_0 = \frac{k_{-2}}{k_2}$, we had: $$D = \dfrac{1}{1+\frac{a^2}{a_0}+\frac{r^4}{r_0}}$$ $$D_1 = \dfrac{a^2}{a_0(1+\frac{a^2}{a_0}+\frac{r^4}{r_0})}$$ $$D_2 = \dfrac{r^4}{r_0(1+\frac{a^2}{a_0}+\frac{r^4}{r_0})}$$ What's worth mentioning is that both $a_0$ and $r_0$ are constant that are related to IPTG(mM) and Arabinose(%). $$a_0 = \dfrac{(6.25+ara^2)(1+\frac{iptg^2}{3.24})}{100ara^2}$$ $$r_0 = \dfrac{1}{2000000(\dfrac{0.19}{1+{\frac{iptg}{0.035}}^2+0.01})}$$ During the transcription process, we had: $$D_1\xrightarrow{k_3}R_{a/r}$$ $$D_2\xrightarrow{k_4}R_{a/r}$$ $$D\xrightarrow{k_5}R_{a/r}$$ where $R_{a/r}$denotes mRNAs of either AraC or LacI. $k_3$, $k_4$ and $k_5$ are transcriptional reaction rates constants. For convenience, we assumed that transcription rate for either AraC or LacI are exactly the same. During the translation and folding processes, we have: $$R_a\xrightarrow{t_a}a_{uf}$$ $$R_r\xrightarrow{t_r}r_{uf}$$ $$a_{uf}\xrightarrow{k_{fa}}a$$ $$r_{uf}\xrightarrow{k_{fr}}r$$ where $a_{uf}$ and $r_{uf}$ are unfolded proteins, $t_a$ and $t_r$ are translational reaction rate constants while $k_{fa}$,$k_{fr}$ are folding rates constants. In degradation process, we had: $$R_{a/r}\xrightarrow{d_{a/r}}\varnothing$$ $$a_{uf}\xrightarrow{\lambda f(x)}\varnothing$$ $$r_{uf}\xrightarrow{f(x)}\varnothing$$ $$a\xrightarrow{\lambda f(x)}\varnothing$$ $$r\xrightarrow{f(x)} \varnothing $$ where $d_{a/r}$, $f(x)$ and $\lambda f(x)$ are degradation rate constants.
According to law of mass reaction, we had: $$\frac{dR_a}{dt} = copy_a(k_3D_1+k_4D_2+k_5D)-d_{a/r}R_a$$ $$\frac{dR_r}{dt} = copy_r(k_3D_1+k_4D_2+k_5D)-d_{a/r}R_r$$ $$\frac{da_{uf}}{dt} = t_aR_a-k_{fa}a_{uf} - \lambda f(x)a_{uf}$$ $$\frac{dr_{uf}}{dt} = t_rR_r-k_{fr}r_{uf} - f(x)r_{uf}$$ $$\frac{da}{dt} = k_{fa}a_{uf} - \lambda f(x)a$$ $$\frac{dr}{dt} = k_{fr}r_{uf} - f(x)r$$ where $copy_a$ and $copy_r$ are plasmid copies that are transfected into E.coli.
Transcriptional and translational processes of genes take time and consequently, protein that combined to promoters can be seen as those started transcription process before a specific time interval $\tau$. Thus we converted three ODEs into DDEs: $$D = \dfrac{1}{1+\frac{a(t-\tau)^2}{a_0}+\frac{r(t-\tau)^4}{r_0}}$$ $$D_1 = \dfrac{a(t-\tau)^2}{a_0(1+\frac{a(t-\tau)^2}{a_0}+\frac{r(t-\tau)^4}{r_0})}$$ $$D_2 = \dfrac{r(t-\tau)^4}{r_0(1+\frac{a(t-\tau)^2}{a_0}+\frac{r(t-\tau)^4}{r_0})}$$
| Parameters and Variables | Meaning | Value |
|---|---|---|
| $a_0$ | Dissociation rate constant of AraC binding and unbinding with promoters | $\dfrac{(6.25+ara^2)(1+\frac{IPTG^2}{3.24})}{101ara^2}$ |
| $r_0$ | Dissociation rate constant of LacI binding and unbinding with promoters | $\dfrac{1}{2000000\frac{0.19}{1.0+(\frac{IPTG}{0.035})^2+0.01}}$ |
| $D$ | Ratio of promoters which don't combine with any protein among all promoters | $\dfrac{1}{1+\frac{a^2}{a_0}+\frac{r^4}{r_0}}$ |
| $D_1$ | Ratio of operons combined with AraC dimer | $\dfrac{a^2}{a_0(1+\frac{a^2}{a_0}+\frac{r^4}{r_0})}$ |
| $D_2$ | Ratio of operons combined with tetrameric LacI | $\dfrac{r^4}{r_0(1+\frac{a^2}{a_0}+\frac{r^4}{r_0})}$ |
| $a$ | AraC protein(activator) | |
| $r$ | LacI protein(repressor) | |
| $R_a$ | mRNA of AraC | |
| $R_r$ | mRNA of LacI | |
| $a_{uf}$ | Unfolded AraC | |
| $r_{uf}$ | Unfolded LacI | |
| $copy_a$ | plasmid containing araC copies that are transfected into E.coli | 50 |
| $copy_r$ | plasmid containing lacI copies that are transfected into E.coli | 25 |
| $k_3$ | Transcriptional reaction rates constants of $D_1$ | 196/min |
| $k_4$ | Transcriptional reaction rates constants of $D_2$ | 0/min |
| $k_5$ | Transcriptional reaction rates constants of D | 5.6/min |
| $d_{a/r}$ | Degradation rate constants | 10.54/min |
| $t_a$ | Translational reaction rate constants of araC | 90/min |
| $t_r$ | Translational reaction rate constants of lacI | 90/min |
| $k_{fa}$ | Folding rate constants of AraC | 0.9/min |
| $k_{fr}$ | Folding rate constants of LacI | 0.9/min |
| $f(x)$ | Degradation rate constants | $\frac{1080}{0.1+X}/min$ |
| $\lambda f(x)$ | Degradation rate constants | $\frac{2887.92}{0.1+X}/min$ |
| $\tau$ | Time delay because of the transcription and translation time | 2min |
Results
We solved these DDEs with R language. To make it more realistic, we also simulated the situation in which lag obeys a specific Gaussian distribution, and the lag $\tau$ changes in every certain interval. The results are below.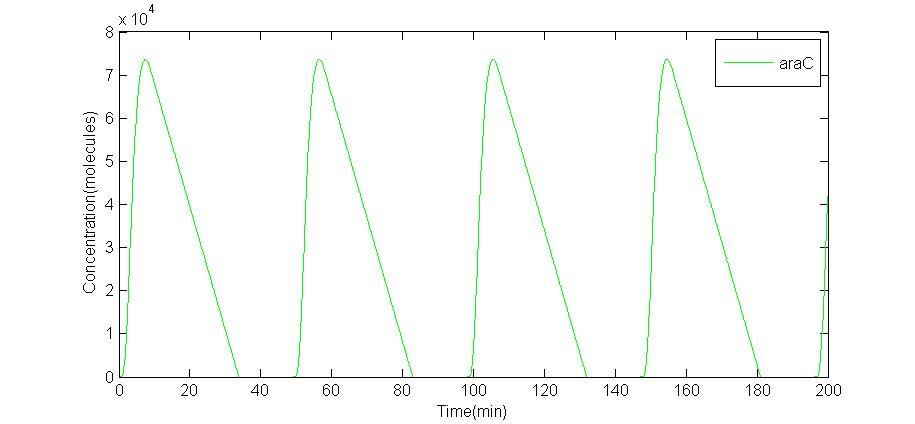
(a) A numeric solve of AraC
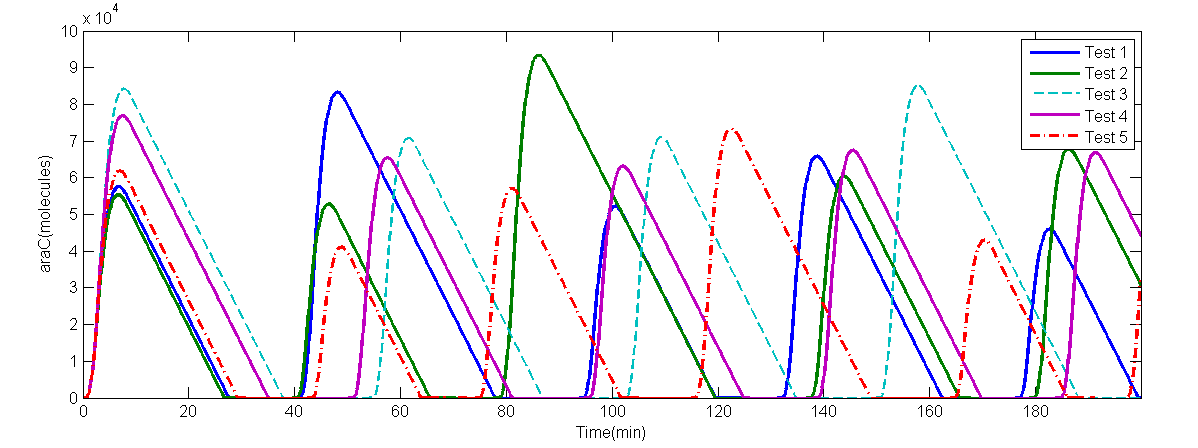
(b) 5 random tests numeric solve of AraC
Fig 1.(a)A numeric solve of AraC when lag $\tau = 2$min, Arabinose concentration is 5%, IPTG concentration is 1mM, time interval is 0.1min. (b)numeric solve of AraC concentration versus time of 5 random tests, when Arabinose concentration is 0.7%, IPTG concentration is 10mM, and $\tau \sim (2.0,0.3^2)$.
The period of this particular solve is 49.0minutes. The numeric solve of DDEs shows that the supposed oscillator is feasible. On the other hand, interval between every adjacent peak is different in a single random test, thus period is calculated by average intervals. Even so, the average period of each random test is different from each other: $T_1$= 43.95min, $T2$= 47.65min, $T_3$= 40.625min, $T_4$ = 39.375min, $T_5$ = 45.975min. Also, the amplitude of each curve is different. The random solve suggests that extern factors might be introduced to force the period to be the same.
References
Team:NTU-Singapore/Modelling/ParameterJesse Stricker et al., 2008, Supplementary Information From A fast, robust and tunable synthetic gene oscillator, Nature 456, 516-519
Acknowledgement


CopyRight ©2013.igem.org All Rights Reserved!
Powered by the iGEM team comes from HuaZhong University of Science and Techology
HUST-China