 "
"
Team:XMU Software
From 2013.igem.org
Revision as of 05:14, 14 September 2013 by YuezhenChen (Talk | contribs)
<!DOCTYPE HTML>






XMU-Software-2013 team consists of 11team members, 2 instructors and 3 advisors. This extraordinarily energetic and creative team is tightly connected by trust, collaboration and affection among its members. Learn more about our team on team page.
Our project includes 2 independent software tools-the brick worker and E’NOTE. The former is a software suit for the evaluation and optimization of biobricks, i.e., promoter, RBS, protein coding sequence and terminator. E’NOTE is a web application serveing as an assistant for experiments. It’s useful functions such as experiments recording and experimental template customization make experimental process easier and more enjoyable. To learn more about our project, please visit Project page.
See more about XMU-Software-2013 safety form on safetyform page
XMU-Software-2013 has hosted a series of activities aiming at facilitating communication and collaboration among iGEM teams, propagating iGEM competition as well as promoting the development of synthetic biology. And these efforts are well paid off and have received satisfying feedback. More information is available at Outreach page.
See more about XMU-Software-2013 Notebook on Notebook page.












TEAM
XMU-Software-2013 team consists of 11team members, 2 instructors and 3 advisors. This extraordinarily energetic and creative team is tightly connected by trust, collaboration and affection among its members.

OUR TEAM
01
Members






















Members

































Instructors

Prof. Bai-shan Fang
Dr. Bai-shan Fang is the professor in College of Chemistry & Chemical Engineering at Xiamen University. In the group of biocatalysis and biotransformation, his research mainly focuses on synthetic biology, mining and the transformation of the enzyme, construction of bio-molecular machines, the application of new biocatalysts. His major role of XMU iGEM team is to mentor for all and to enlighten the members.

Associate Prof. I-Son Ng
Dr. I-Son Ng is the associate professor in College of Chemistry & Chemical Engineering at Xiamen University. Her research interests are biofuel, engineering of enzyme and protein, zymology, genetic engineering, biochemical separation procedures and proteomics. Her role for the project is to provide suggestions and instruments.
Advisors

Ruosang Qiu
It is great for a team to have an omniscient advisor, even better an inspiring one. Ruosang Qiu , our beloved advisor, definitely is offering both. Her hard work as well as undoubted adorable personality is the motivation of our team members’ efforts, her clear mind combined with provident planning lays the foundation of our successful project, To quote her words: I’ was a happy iGEMer in 2012, I’m going to make you all happy iGEMers in 2013.

Xin Wu
The past three years have seen his tremendous dedication to iGEM, Xin Wu, a passionate team member in XMU-China-2011, a devoted team leader in XMU-China-2012 and now, an invaluable advisor in XMU-Software-2013. Had it not been Xin Wu's constant encouragement and guidance, we,the inexperienced iGEMers might have been faint-hearted and failed to face up to the challenges during the process. It is drawing on his expertise in synthetic biology and proficiency in iGEM competition that we have solved the seemingly unsolvable and conquered the seemingly unconquerable.

Youbin Mo
Youbin Mo is one of the great advisors of XMU software team in 2013. As a computational biophysicist, y, he is an unquestionable master of biological model and computer programing. In the meanwhile, website constructing is also Mo’s technical ability which he acquired by participating the iGEM last year. Youbin gives play to his talent by teaching fundamental program skills to new iGEMers as well as directing them to be self-reliant synthetic biologists.
ACKNOWLEDGEMENT
02
All work described on this wiki or on our parts registry pages was done by iGEM Team XMU-China 2012. We managed the whole project, from planning to financing the complete lab work by ourselves. Nevertheless we could not have done all this work without the help, advice and guidance of several people. Therefore, special thanks to the following people:
Our advisor Graduate Xuesong Zheng for helping us in fluorescence test and immobilization.
Prof. Xiaodong Chen for providing us many instruments such as ELISA reader to test the fluorescence and his students Song Huang, Renpan Deng, Xiang You for experimental guidance.
Prof. Yinghua Lu and his student Shiduo Zhang for helping us make the PDMDAAC-NaCS microcapsules.
Dr. Ying Zeng, Kunming Institute of Botany, Chinese Academy of Sciences, who has provided us with valuable guidance in constructing gene circuits.
OUR CITY&UNIVERSITY
03
Xiamen
Xiamen, also known as Amoy to the west, is a cozy city located in the southeastern part of China, and has a relaxing coastal charm with a population of 1.3 million. It's a historical harbor city which was founded in the mid-14 century, in the early years of the Ming Dynasty. In the early 1980's, Xiamen was declared as one of China's first Special Economic Zone, taking advantages of the city's heritage as a trading center and the proximity to Taiwan. In 2004 the city won the finals of the world's Human Settlements and Environment Award, "Nations in Bloom". Xiamen is one of China's most attractive and best-maintained resort city, and attracts a large number of foreign and local tourists. The city is easily accessible by air, and there are direct flights from Hong Kong, Kuala Lumpur, Osaka, Seoul, Singapore and Tokyo. Within China, Xiamen airport is linked to more than 30 domestic airports.


Xiamen University
Xiamen University (XMU), also known as Universitas Amoiensis in Latin, is one of the top universities in China. It was founded in 1921 by Tan Kah-Kee, the well-known patriotic overseas Chinese leader. As an integrated university, XMU owns a comprehensive branches of discipline as well as many specialized institutes. Economy, counting, chemistry, life science and marine science all win high fame nationwide and even worldwide. The main campus of XMU locates in a picturesque setting between the sea and a scenic mountain, spreading over 150 hectares, and is generally regarded as the most beautiful campus in China.
PROJECT
Our project includes 2 independent software tools-the brick worker and E’NOTE. The former is a software suit for the evaluation and optimization of biobricks, i.e., promoter, RBS, protein coding sequence and terminator. E’NOTE is a web application serveing as an assistant for experiments. It’s useful functions such as experiments recording and experimental template customization make experimental process easier and more enjoyable. To learn more about our project, please visit Project page.
Terminator
03
Background
Gene expression in both prokaryotes and eukaryotes is frequently controlled at the level of transcription. This process can be represented as a cycle consisting of four major steps: (1) promoter binding; (2) RNA chain initiation; (3) RNA chain elongation; and (4) termination. Since regulatory controls are exerted at each step, an understanding of the mechanism of each step is of general importance in understanding gene expression.
In the promoter part of our program, we’ve discussed the mechanism of promoter binding step and how it affects the transcription level. To complete our biobrick evaluation program and to better understanding of transcription process, we integrated the software developed by 2012 iGEM team SUSTC-Shenzhen-B to realize the prediction of transcription termination efficiency.
Introduction
Termination, the last step of the transcription cycle, occurs when the RNA polymerase releases the RNA transcript and dissociates from the DNA template. It is important that transcription is imperfectly terminated at some terminator so that the ratio of the amount of the mRNA transcribed from upstream and that from downstream of the terminator is controlled. This regulation is quantified by the termination efficiency (%T),
Two mechanisms of transcription termination and two classes of termination signals have been described in bacteria: rho-dependent and rho-independent.
Rho-independent (also known as intrinsic) terminators are sequence motifs found in many prokaryotes that cause the transcription of DNA to RNA to stop. These termination signals typically consist of a short, often GC-rich hairpin followed by a sequence enriched in thymine residues.
The conventional model of transcriptional termination is that the stem loop causes RNA polymerase to pause and transcription of the poly-A tail causes the RNA: DNA duplex to unwind and dissociate from RNA polymerase.
Algorithm
In 2011, iGEM team SUSTC-Shenzhen-B developed a software tool TTEC to predict terminator efficiency. It takes DNA sequences as input and returns the terminator efficiency value.
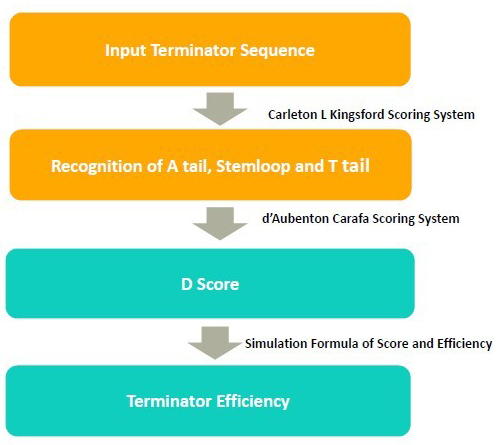
In the algorithm, it takes 3steps to calculate the terminator efficiency:
1. Use RNA folding algorithm to predict the secondary structure of terminator and and recognize A tail, stemloop and T tail.
2. From the secondary structure, we calculate the free energy of stem loop, and generate a score by considering stem loop free energy and T tail
3. From the score, we predict the terminator efficiency based on the score-terminator equation.
The prediction of secondary and recognition of A tail, stemloop and T tail are achieved by Kings ford scoring system.
Kingsford Scoring System
In 2007, Carleton L Kingsford et al described TransTermHP1, a new computational method to rapidly and accurately detect Rho-independent transcription terminators.
They put forward an algorithm to predict Rho-independent terminators.The first 15 bases of the potential tail sequence are scored using a function:

where

for n=1...15 and =1.
The energy of potential hairpin configurations adjacent to a reference position can be found efficiently with a dynamic programming algorithm. The table entry hairpin_score[i,j] gives the cost of the best hairpin structure for which the base of the 5' stem is at nucleotide position i and the base of the 3' stem is at position j. The entry hairpin_score[i,j] can be computed recursively as follows:

The function energy(i,j) gives the cost of pairing the nucleotide at i with that at j, and loop_pen(n) gives the cost of a hairpin loop of length n. The hairpin's loop is forced to have a length between 3 and 13 nt, inclusive, by setting loop_pen(n) to a large constant for any n outside that range. The constant 'gap' gives the cost of not pairing a base with some base on the opposite stem and thus introducing a gap on one side of the hairpin stem.
Table 1
Parameters used to evaluate hairpins
Pairing Energy
G-C -2.3
A-T -0.9
G-T 1.3
Mismatch 3.5
Gap 6.0
Loop_pen(n) 1•(n - 2)
Parameters used to evaluate the energy of a potential hairpin where n is the length of the hairpin loop
The D score is calculated by Carafa Scoring System
Carafa Scoring System
Scoring System 2 is based on the model created by d'Aubenton Carafa2 .The score of terminator consists of two parts, the free energy of stemloop and the score of 15 nt poly T tail. The free energy of stemloop is calculated using Loop Dependent Energy Rules3. The minimization of the free energy also determined the secondary structure of the stemloop. T tail score is calculated by the formula given by d’ Aubenton Carafa.
Detailed Calculation of Score
1. Some definitions3
i. Closing Base Pair
For an RNA sequence, we number it from 5’ to 3’ . If i < j and nucleotides ri and rj form a base pair,we denote it by i.j. We call base ri’ or base pair i’.j’ is accessible from i.j if i <i’ ( <j’ ) <j and if there is no other base pair k.l so that i <k <i’ ( <j’ ) <l <j. We denote the collection of base and base pair accessible from i.j by L(i,j). Then i.j is the closing base pair. Here “L” means loop.
ii. n-loop
If the loop contain n – 1 base pairs, we denote it by n-loop. (Because there is a closing base pair, so we denote it by n-loop even though the closing base pair is not included in the loop.)
Here we can divide loops which may be formed in the terminator secondary structure into two kinds.
1-loop : Hairpin loop(size of loop shouldn’t be smaller than 3)
2-loop : Interior Loop(right strand size and left strand size are both bigger than 0.)
Buldge(Size of one strand is bigger than 0 and that of another strand is 0.)Stack(size of the loop is 0.)
2. Calculation of the Minimum Free Energy Change of Stemloop Formation4 Assume i.j is the closing base pair of the loop
G(i,j)= min { GH ( i , j ) , GS( i , j ) + G ( i + 1 , j – 1 ) , GBI( i , j ) } ;
GBI ( i , j ) = min{ gbi( i , j , k , l ) + G( k , l ) } for all 0 < k – i + l – j - 2 < max_size
G(i,j) is the minimum free energy change of stemloop formation. GH is the free energy change to form a hairpin loop. GS is the free energy change to form a stack. GBI is to calculate the minimum free energy change of structure containing 2-loop. gbi(i,j,k,l) is the free energy change to form 2-loop.
3.Calculation of T Tail Score
Here we consider 15 nucleotide in the downstream of stemloop. T tail score nT is calculated as follows :

In our program, if the length of the T tail( n ) is less than 15, we will only consider n nucleotides. If TL is more than 15, we will only consider 15 nucleotides.
4.Calculation of Score
Score = nT * 18.16 + deltaG / LH * 96.59 – 116.87
Here nT is T tail score. deltaG is the minimum free energy change of stemloop formation. LH is the length of stemloop5.6
References
1. Kingsford, C. L.; Ayanbule, K.; Salzberg, S. L., Rapid, accurate, computational discovery of Rho-independent transcription terminators illuminates their relationship to DNA uptake. Genome biology2007, 8 (2), R22.
2. Carafa, Y. d. A.; Brody, E.; Thermes, C., Prediction of rho-independent Escherichia coli transcription terminators: A statistical analysis of their RNA stem-loop structures. Journal of molecular biology 1990,216 (4), 835-858.
3. Manual of Mfold Version 3.5.
4. http://unafold.math.rpi.edu/lectures/old_RNAfold/node2.html.
5. Lesnik, E. A.; Sampath, R.; Levene, H. B.; Henderson, T. J.; McNeil, J. A.; Ecker, D. J., Prediction of rho-independent transcriptional terminators in Escherichia coli. Nucleic acids research 3583-3594.
6. Sugimoto, N.; Nakano, S.-i.; Katoh, M.; Matsumura, A.; Nakamuta, H.; Ohmichi, T.; Yoneyama, M.; Sasaki, M., Thermodynamic parameters to predict stability of RNA/DNA hybrid duplexes. Biochemistry 1995, 34 (35), 11211-11216.
RBS-decoder
02
Abstract
RBS-decoder is a software tool for evaluating the relative strength of RBS sequence. This program use the same method—PWM to calculate the similarity between RBS sequence and the position frequency matrix of SD sequence and transform the similarity to the relative strength of an RBS sequence.
Background
Translational efficiency in Escherichia coli is generally determined at the stage of initiation. There are several principal mRNA sequence elements which can affect the kinetics of ternary initiation complex formation (30 S-mRNA-fMet-tRNA): the initiation codon, the Shine-Dalgarno (SD) sequence, the spacing between those two elements, and other factors.
Introduction
How do baterial Ribosome Binding Sites work?
The bacterial ribosome binds to particular sequences on an mRNA, primarily the Ribosome Binding Site (RBS) and the start codon (ATG). The RBS base-pairs with an RNA molecule that forms part of the bacterial ribosome (the 16s rRNA), while the start codon base-pairs with the initiator tRNA which is bound to the ribosome. In addition to the sequences of the RBS and the start codon being important, these two sequences need to be positioned approximately 6-7 nucleotides apart so they can both make contact with the appropriate parts of the ribosome complex1.
The Shine-Dalgarno Sequence

The RBS sequence logo representing the sequences of 149 RBS from E. coli. The height of each letter represents the frequency of the base at that location. From Tom Schneider, "A Gallery of Sequence Logos".
The end of the 16s rRNA that is free to bind with the mRNA includes the sequence 5′–ACCUCC–3′. The complementary sequence, 5′–GGAGGU–3′, named the Shine-Dalgarno sequence, can be found in whole or in part in many bacterial mRNA. Very roughly speaking, ribosome binding sites with purine-rich sequences (A's and G's close to the Shine-Dalgarno sequence will lead to high rates of translation initiation whereas sequences that are very different from the Shine-Dalgarno sequence will lead to low or negligible translation rates. The sequence is named after the researchers who discovered the high frequency of the sequence upstream of bacterial coding sequences . You can see how common the sequence is by looking at the RBS sequence logo on the right (where the height of a letter indicates the frequency of the letter at that location).
Algorithms
As we know, the RBS strength is greatly influenced by the SD sequence, where the 16s RNA of the ribosom binds to, so the strength can be determined by the free binding energy between the SD sequence and the 16s RNA. So we designed a program calculating the free binding energy but the results show that the correlation between the free energy and the strength of RBS is rather weak (R2=0.5517). So we decide to find other algorithms for better accuracy.
Inspired by the strength prediction algorithms used in promoter part, in which the similarity to the sigma factors’ PWM is interlocking with the binding affinity between the protein and DNA sequence. We obtained the Position Frequency Matrix of SD sequence of E.coli and use the PMW method (illustrated in detail in the promoter part) to calculate the similarity between the RBS sequence and the Position frequency sequence, what is different from the promoter is that, the spacer length between the SD sequence and the startcodon and the start codon itself both act as constraints in locating the SD sequence, which is confined to 3-16 bp and ATG/TTG/GTG. And similar to the prediction of promoter strength, the spacer length between the SD sequence also contribute to the RBS strength, the weight of the influence of the spacer on strength is determined by our experiments in which we either mutated the SD sequence in a RBS sequence or change the spacer length to figure out the relative influence of these two factors 2.

Results
We use the RBS sequence listed on the iGEM registry with experimentally determined relative strength, and the correlation between the RBS strength predicted by our software and the actual relative strength is strong, with a determination coefficient value 0.8039.

Future work
Due to scarcity of experimental data, the relative weight of the SD sequence and the spacer length used currently is roughly determined which may undermine the accuracy of RBS strength prediction. For further improvement of our program, we’ll try to obtain more reliable experimental data to accurately determine the weight used in our algorithm and hopefully elevate the accuracy of RBS-decoder.
In the next version of RBS-decoder, the secondary structure of the RBS sequence will be shown on the software and we’ll also include the other species’ SD sequence data in order to analyze predict the strength of RBS.
Reference
1. Ma, J.; Campbell, A.; Karlin, S., Correlations between Shine-Dalgarno sequences and gene features such as predicted expression levels and operon structures. Journal of bacteriology 2002, 184 (20), 5733-5745.
2. Noguchi, H.; Taniguchi, T.; Itoh, T., MetaGeneAnnotator: detecting species-specific patterns of ribosomal binding site for precise gene prediction in anonymous prokaryotic and phage genomes. DNA research 2008, 15 (6), 387-396.
Promoter-decoder
01
Abstract
Pro-decoder is part of our software designed to analyze and evaluate promoters. The software use PWM method to calculate the similarity between promoter sequences and the position frequency matrix of transcription factor binding sites (TFBS) to locate the TFBS as well as predict the relative strength of the promoter. So the user may have a better understanding of the promoter’s regulation mechanism and the key sites which influences the promoter’s performance mostly.
Background
Sigma factors
Bacteria encode several thousands of different proteins, which are necessary for normal cell functions or for adaptation to environmental changes. These proteins are not required at the same time or in the same amount. Regulation of gene expression therefore enables the cell to control the production of proteins needed for its life cycle or for adaptation to extracellular changes. This regulation in turn makes it possible for the bacterium to adequately adapt to rapid changes in the environment. The various steps during transcription and translation are therefore subject to different regulatory mechanisms. The most prominent step in gene regulation is the initiation of transcription in which the DNA-dependent RNA polymerase (RNAP) is the key enzyme. The RNAP or the RNAP core enzyme is the catalytic machinery for the synthesis of RNA from a DNA template. However, RNAP cannot initiate transcription by itself. Initiation of transcription requires an additional polypeptide known as a sigma-factor. Sigma-factors are a family of relatively small proteins that can associate in a reversible way with the RNAP core enzyme. Together, the sigma-factor and the RNAP core enzyme form an initiation-specific enzyme, the RNAP holoenzyme

The sigma-factor directs RNA polymerase to a specific class of promoter sequences. Most bacterial species synthesize several different sigma-factors that recognize different consensus sequences. This variety in sigma-factors provides bacteria with the opportunity to maintain basal gene expression as well as for regulation of gene expression in response to altered environmental or developmental signals.
The frequency at which the RNAP holoenzyme initiates transcription, also known as the strength of a promoter, is influenced by the promoter sequence and the conformation of the DNA in the promoter region. The sigma-factors recognize two conserved sequences in the promoter region, known as the promoter consensus sequence. Sigma-factors or fragments of sigma-factors bind specifically to promoter DNA sequence and by specific base pair and amino acid substitutions in the promoter consensus sequences or sigma factors. Most bacterial species synthesize several different sigma-factors which direct the RNAP holoenzyme to distinct classes of promoters with a different consensus sequence. This variety in sigma-factors provides the bacterium with the opportunity to maintain basal gene expression as well as for regulation of gene expression in response to specific environmental stimuli.
The identification of bacterial promoters is an essential step in the elucidation of gene regulation. As a general rule, the more complex the life-cycle and environmental niche of a bacterium, the greater the number of sigma factors with corresponding promoter types. Typically however, the most common promoter type is that which regulates the housekeeping genes and the corresponding major sigma-factor is shared by all bacteria (sigma 70 in the well studied Escherichia coli, and its homologues in other species). The binding site for the sigma70-family of promoters is defined by two consensus hexamers, TTGACA and TATAAT, located at approximately −35 and −10, respectively relative to the transcript start site (TSS) and spaced 15–21 base pairs (bp) apart2. RNA polymerase core enzyme associates with the major sigma-factor to form the holoenzyme which in turn binds to its cognate promoters to initiate transcription.


In prokaryotes, the minimum requirement for RNA polymerase binding is recognition of the promoter by the sigma factor. In general, prokaryotic RNA polymerases can interchange a number of sigma factors which bind and initiate different groups of genes3.
Transcription Factors

Sigma factors are essential for the transcription initiation in Escherichia coli. In addition, promoter strengths are not determined purely by the binding of the sigma factor. other transcription factors can bind specific sequences surrounding or overlapping the promoter to either activate or repress transcription4. The mechanism is Transcriptional activators and repressors contribute to and detract from the accessibility of DNA by the RNA polymerase. These transcription-regulating nuclear proteins bind to specific binding sites in the regulatory regions (e.g. promoters, enhancers) of the genes thus providing their activation or repression.


Computational methods of predicting TF binding sites in DNA are very important for understanding the molecular mechanisms of gene regulation.
The binding sites of the same transcription factor show a significant sequence conservation, which is often summarized as a short (5–20 bases long) common pattern called a transcription factor binding site (TFBS) or binding consensus. Our software aims to figure out the possible TFBS in promoters and precisely locate the TFBS so that the user may know the exact sites that play a role in regulating the transcription.
In prokaryotes (lower organisms without nuclei), there are fewer TFs, their motifs tend to be relatively long and the strength of regulation for a particular gene often depends on how closely a particular site matches the consensus for the motif. The more mismatches to the consensus in a binding site, the less often the TF will bind and therefore the less control it will exert on the target gene. So our software will calculate the similarity between the possible TFBS in the promoter and the standard motifs so the user will know to which extent the transcription factor will control the promoter transcription
Primer design
To facilitate the design of PCR primers of various promoters, we’ve developed an additional function, namely, primer design in this part og our program. After inputting the promoter sequence, the software will figure out the most suitable primers based on the theory of Thomas K¨ampke1, Markus Kieninger, and Michael Mecklenburg.
Data source
RegulonDB
Genes and operons that are under control of the same TF are members of that TF’s regulon. Although methods for the prediction of regulons have been substantially improved, they are still far from perfect.
Comparative genomics tools can be used to predict regulons in bacterial genomes but the procedure can lead to incorrect regulon calling. Despite this drawback, several regulon databases are available that are based on comparative genomics methods and lack experimental evidence.
Probably the extended and accurate databases of regulons for E.coli are RegulonDB which provides the data source for our program.
Algorithm
Experimental results show that these are the strongest promoters that have been characterized in vitro so far and confirm the hypothesis that the consensus promoter sequence is “best.” To calculate the similarity between the promoter sequence and the best sequence, we implement the PWM method 5.
PWM (Position Weight Matrix)
Molecular techniques for the identification of promoters are both costly and time consuming, hence in silico methods are an attractive and well explored alternative. The most common in silico method to identify _70 promoters uses position weight matrices (PWMs) and depends on the relative conservation of the transcription factor binding site (TFBS, or motifs ).
The algorithm can be divided into two parts regarding to the difference between the motifs of sigma factors and other transcription factors.

Part 1: the recognition of other transcription factors6
Other transcription factors are proteins that can bind to a specific DNA sequence (motifs) and regulate the promoter’s transcription. To recognize these possible motifs in a given promoter sequence, we calculate the Matrix Similarity Score (MSS) of every possible sites in the promoter sequence using the position frequency matrix of 86 transcription factors published by RegulonDB. The algorithm reports only those matches of a matrix that have got MSS higher than the settled threshold. And MSS for a subsequence x of the length L is calculated in following steps:


fi,B, frequency of nucleotide B to occur at the position i of the matrix (B ∈{A, T, G, C})

fimin, frequency of the nucleotide which is rarest in position i in the matrix

fimax, highest frequency in position i.
The information vector

describes the conservation of the positions i in a matrix (5).Multiplication of the frequencies with the information vector leads to a higher acceptance of mismatches in less conserved regions, whereas mismatches in highly conserved regions are very much discouraged. This leads to a better performance in recognition of TF binding sites if compared with methods that do not use the information vector.
To determine the best threshold of the motif finding algorithm, we test various threshold values and analyze the true negative and false positive rate of each threshold value. The ideal threshold is supposed to have both the least true negative and false positive rates.

The picture above shows part of our test results and to keep both the true negative and false positive rates at a reasonable level, we adopt 3 threshold values, namely, low (0.5977) , median (0.0778) and high (0.85), with a true negative rate at 0.1, 0.5, and 0.7 respectively. And for more flexibility, we also allow the users to set their own thresholds.
Part2: the recognition of sigma factor motif and the evaluation of relative promoter strength.
In the case of sigma 70 factors, the motifs are the −35 and −10 hexamers. enclosing a spacer of length 15–19 bp.
Given a known or predicted TSS location, the corresponding predictions for the −10 and −35 hexamers are located using a combination of two PWMs which are derived from literatures. For any known or putative TSS, the −35 and −10 hexamers are located upstream of the TSS by searching for the highest combination of PWM scores, subject to two constraints:
(i) That the spacer length (the number of base pairs between the −35 hexamer and the −10 hexamer) should lie in the range {14–20};

(ii) the total MSS (our results are the sum of the scores for the −10 and −35 hexamers and therefore lie in the interval [0,2], with a score of 2 corresponding to the joint consensus TTGACA (−35) and TATAAT (−10).

In prokaryotes, the strength of sigma factor regulation for a particular gene often depends on how closely a particular site matches the consensus for the motif. The more mismatches to the consensus in a binding site, the less often the sigma factor will bind and therefore the less strength the promoter will have. Experiments have confirmed the hypothesis that the consensus promoter sequence is “best”. We set the best promoter strength to 100% and calculate the relative strength of a given promoter by the Score (promoter)
Primer design
A primer pair (p, q) is assigned the scoring vector
sc(p, q) = (|p|, |q|,GC(p),GC(q), Tm(p), Tm(q), sa(p),sa(q), sea(p), sea(q), pa(p, q), pea(p, q))T ∈ R12
All primers are designed to have ideal values of length,GC content, and melting temperature which are specified externally by the designer of the hybridization experiment. These ideal values are to be specified for forward and reverse primers. The ideal score vector or reference vector for the primer pair is
                              
scideal =(lengthf , lengthr,GCf ,GCr , Tm,f ,Tm,r , 0, 0, 0, 0, 0, 0)T.
All ideal annealing values are set to zero and typically
Tmf= Tm,ras well as GCf = GCr . The final assessment of a primer pair (p, q) can be its deviation from the reference in terms of the l1-distance

Here, we employ a weighted distance

with weights given in the following table.

The formulas for calculating the variations above are provided in 7
Results
Sigma factors recognition
To test the ability of our program to recognize the exact type of promoters, i.e., to nail down by which sigma factor a promoter is regulated, we run our program with 100 various promoters sequences whose type have already confirmed experimentally. And we’ve recognized 56 of them correctly. The results are showed below.
TFBS Location
Specifically, we use the sigma70 promoter sequence with annotated -35 and -10 region provided by RegulonDB to test the correct prediction rate of the binding site of a specific transcription. We input 89 sigma70 promoter sequences and run our program to precisely locate the sigma factor binding site. Results show that the correct site prediction rate of our program is 64%.
The test results are as follows. the numbers represent the site of actual-35 motif, the actual spacer length, the predicted site and predicted spacer length respectively.
Promoter strength correlation
Making an attempt to predict the promoter strength through motif similarity and spacer length is one of our software’s shine points. We input XX promoter sequences with experimentally determined strength data. And the result shows that the determination coefficient is X.
Future work
Apply our algorithms to more species. Now Pro-decoder is designed expressly for prediction and evaluation of E.coli promoters, in the near future we’ll study the transcription regulation mechanism of other species and try to apply our algorithms to a extended range of species.
Enhance promoter strength prediction accuracy. Because our experimental data is so limited, the weight of the spacer length and the motif similarity is roughly determined, which lead to a weak correlation between the promoter strength. In the future we hope to obtain more experimental data with regard to the effect of spacer length and motif similarity having on promoter strength so we can revise the weight coefficient of the two factors and get more reliable results.
The next version of this part of our program will be able to analyze not only the promoters of E.coli, but other species such as Bacillus subtilis, we’ll integrate the transcription factor binding site data of more species into our database and use PWM algorithm to predict the TFBS in the promoters.
References
1. Wösten, M., Eubacterial sigma‐factors. FEMS microbiology reviews 1998, 22 (3), 127-150.
2. Shultzaberger, R. K.; Chen, Z.; Lewis, K. A.; Schneider, T. D., Anatomy of Escherichia coli σ70 promoters. Nucleic acids research 2007, 35 (3), 771-788.
3. Paget, M.; Helmann, J. D., The sigma70 family of sigma factors. Genome Biol 2003, 4 (1), 203.
4. Jensen, S. T.; Liu, X. S.; Zhou, Q.; Liu, J. S., Computational discovery of gene regulatory binding motifs: a Bayesian perspective. Statistical Science 2004, 19 (1), 188-204.
5. (a) Rhodius, V. A.; Mutalik, V. K., Predicting strength and function for promoters of the Escherichia coli alternative sigma factor, σE. Proceedings of the National Academy of Sciences 2010, 107 (7), 2854-2859; (b) Mulligan, M. E.; Brosius, J.; McClure, W. R., Characterization in vitro of the effect of spacer length on the activity of Escherichia coli RNA polymerase at the TAC promoter. Journal of Biological Chemistry 1985, 260 (6), 3529-3538; (c) Qureshi, S. A.; Jackson, S. P., Sequence-Specific DNA Binding by the<i> S. shibatae TFIIB Homolog, TFB, and Its Effect on Promoter Strength. Molecular cell 1998, 1 (3), 389-400.
6. Kel, A. E.; Gößling, E.; Reuter, I.; Cheremushkin, E.; Kel-Margoulis, O. V.; Wingender, E., MATCHTM: a tool for searching transcription factor binding sites in DNA sequences. Nucleic acids research 2003, 31 (13), 3576-3579.
7. Kämpke, T.; Kieninger, M.; Mecklenburg, M., Efficient primer design algorithms. Bioinformatics 2001, 17 (3), 214-225.
SynoProteiner
04
Abstract
Our software team mainly focuses on programming the software by two methods using genetic algorithm, evaluating both optimization of single codon and codon pair and hence determining the fittest optimized sequence for expression in heterologous host cell.
After choosing the method and deciding the host cell, user inputs an original encoding sequence. By substitute the synonymous codons in the sequence, a new sequence is generated. Caluculate the fitness value and choose between the two sequences under the rule that the larger the value is, the higher the possibility of being chosen will be. The chosen sequence will be regarded as the original encoding sequence and repeat the steps until iteration stop criterion. By the way, user can click the “values” button to input the values needed in genetic algorithm (default value: population size—200, hereditary algebra—100, mutation probability--0.1, crossover probability--0.1).
Background
synonymous codons and the efficiency
Except methionine and tryptophan, all amino acids can be encoded by two to six synonymous codons, resulting from the degeneracy of the genetic code.1 However, unequal utilization of the synonymous condons leads to the phenomenon of codon usage bias, which is mainly due to natural selection, mutation and genetic drift.2 According to related studies, codon usage bias has certain connection with gene expression level. 3The larger the value of codon usage bias is, the higher gene expression will be. So the problem, how to substitute the synonymous codons aimed at raising the efficiency of gene expression and thus increasing the production of recombination protein in heterologous host cell, is expected to be addressed.
Introduction
Balance with single codon and codon pair
Individual codon usage optimization has been attached importance to, taking Codon optimizer4, Gene Designer5, OPTIMIZER6 for example. Subsequently, people found the effect of gene expression optimization cannot be perfect just by single codon optimization. Codon pair, namely the pair of k-th and (k+1)-th codons from the 5’ to 3’ end, is another crucial factor. Due to potential tRNA-tRNA steric interaction within the ribosomes7, the usage of rare condon pairs, which correlate with translation elongation, decrease protein translation rates8. Optimization of individual codon has an influence on the corresponding codon pair resulting in maybe-not-the-best codon pair optimization. In the same way, optimizing codon pair merely contributes to maybe-not- the-best single codon optimization. Therefore, it is a challenging way for us to apply a method considering and weighing the effects of single codon and codon pair optimization and thus make the whole best.
Our software team mainly focuses on programming the software by two methods using genetic algorithm, evaluating both optimization of single codon and codon pair and hence determining the fittest optimized sequence for expression in heterologous host cell.
Host Cell
Considering E. coli and S. cerevisiae are the ideal hosts for recombinant proteinexpression, and Gram-positive bacterium L.lactis and methylotrophic yeast P.pastoris are also promising candidates for expressing recombinant proteins9, we attached importance to selecting these four kinds of bacterium as host cell to optimize the sequence.
Algorithm
Part I—method I: Fitness10
Fitness function:

In the function,

cpi is a value larger than zero, ranging from 10-4 to 0.5,fitcp(g) is the fitness function of the codon pair,fitsc(g) is the fitness function of the single codon,w((c(k),c(k+1)) is the weight of codon pairs in sequence g,|g| is the length of encoding sequence,c(k) is k-th codon in the sequence, is the target ratio of k-th codon,
is the target ratio of k-th codon,  is the actual ratio of k-th codon in the sequence,the best value of cpi is 0.2 in the software.
is the actual ratio of k-th codon in the sequence,the best value of cpi is 0.2 in the software.
is the target ratio of k-th codon, is the actual ratio of k-th codon in the sequence,the best value of cpi is 0.2 in the software.In the function, the target ratio of k-th codon can be approximated by the equation below:

In the function, weight can be calculated by the equation below:


 stands for the ratio of single codon ckin the complete genome'
stands for the ratio of single codon ckin the complete genome' is the number of pair (ci,cj) in high-expression genes,and high-expression genes are genes whose copy numbers of mRNA can be detected at least 20 per cell.
is the number of pair (ci,cj) in high-expression genes,and high-expression genes are genes whose copy numbers of mRNA can be detected at least 20 per cell. 
syn(ck)stands for the synonymous codon set related to ck, equals to the number of amino acid encoded by ci in the whole protein set.
equals to the number of amino acid encoded by ci in the whole protein set.
equals to the number of amino acid encoded by ci in the whole protein set. Part II—method II: MOCO11
Basic Table
Based on the table below, we calculate function of single codon (ICU) , function of codon pair (CCO) and function of the combination of two (MOCO). MOCO aims at make the optimization of whole best by calculating the relative effect of ICU and CCO.




Calculation of ICU
max    
s.t.    


Calculation of CC
max    
s.t.    


Calculation of MOCO
The MOCO problem is as follows (NSGA-II algorithm applied):
1. Randomly initialize a population of coding sequences for target protein.
2. Evaluate ICU and CC fitness of each sequence in the population.
3. Group the sequences into nondominated sets and rank the sets.
4. Check termination criterion.
5. If termination criterion is not satisfied, select the “fittest” sequences (top 50% of the population) as the parents for creation of offsprings via recombination and mutation.
6. Combine the parents and offsprings to form a new population.
7. Repeat steps 2 to 5 until termination criterion is satisfied.
The identification and ranking of nondominated sets in step 3 is performed via pair-wise comparison of the sequences’ ICU and CC fitness. For a given pair of sequences with fitness values expressed as  and
and  , the domination status can be evaluated as follows:
, the domination status can be evaluated as follows:
and , the domination status can be evaluated as follows:• If  >
> and
and  >=
>= , sequence 1 dominates sequence 2.
, sequence 1 dominates sequence 2.
> and >= , sequence 1 dominates sequence 2.• If >= and > , sequence 1 dominates sequence 2.
>= and > , sequence 1 dominates sequence 2.• If < and <= , sequence 2 dominates sequence 1.
< and <= , sequence 2 dominates sequence 1.• If <= and < , sequence 2 dominates sequence 1.
<= and < , sequence 2 dominates sequence 1.Future work
1. Shortening the computing time of the software
2. Expanding the range of the host cells
3. Improving bacterium’s resistance to toxic molecule
4. Advancing existed paths of synthetic biology by the method
5. Designing new paths of synthetic biology by the method
6. Increasing the output of recombinant protein
7. Predicting the expression of heterologous gene in a new host cell
References
1. Grantham, R.; Gautier, C.; Gouy, M.; Mercier, R.; Pave, A., Codon catalog usage and the genome hypothesis. Nucleic acids research 1980, 8 (1), 197-197.
2. Hershberg, R.; Petrov, D. A., Selection on codon bias. Annual review of genetics 2008, 42, 287-299.
3. Gouy, M.; Gautier, C., Codon usage in bacteria: correlation with gene expressivity. Nucleic acids research 1982, 10 (22), 7055-7074.
4. Fuglsang, A., Codon optimizer: a freeware tool for codon optimization. Protein expression and purification 2003, 31 (2), 247-249.
5. Villalobos, A.; Ness, J. E.; Gustafsson, C.; Minshull, J.; Govindarajan, S., Gene Designer: a synthetic biology tool for constructing artificial DNA segments. Bmc Bioinformatics 2006, 7 (1), 285.
6. Puigbò, P.; Guzmán, E.; Romeu, A.; Garcia-Vallvé, S., OPTIMIZER: a web server for optimizing the codon usage of DNA sequences. Nucleic acids research 2007, 35 (suppl 2), W126-W131.
7. Smith, D.; Yarus, M., tRNA-tRNA interactions within cellular ribosomes. Proceedings of the National Academy of Sciences 1989, 86 (12), 4397-4401.
8. Coleman, J. R.; Papamichail, D.; Skiena, S.; Futcher, B.; Wimmer, E.; Mueller, S., Virus attenuation by genome-scale changes in codon pair bias. Science 2008, 320 (5884), 1784-1787.
9. (a) Wildt, S.; Gerngross, T. U., The humanization of N-glycosylation pathways in yeast. Nature Reviews Microbiology 2005, 3 (2), 119-128; (b) Morello, E.; Bermudez-Humaran, L.; Llull, D.; Sole, V.; Miraglio, N.; Langella, P.; Poquet, I., Lactococcus lactis, an efficient cell factory for recombinant protein production and secretion. Journal of molecular microbiology and biotechnology 2007, 14 (1-3), 48-58.
10. 帝斯曼知识产权资产管理有限. 公司实现改进的多肽表达的方法: 中国, 200780024670.5[P]. 2009-07-22
11. Chung, B.; Lee, D.-Y., Computational codon optimization of synthetic gene for protein expression. BMC systems biology 2012, 6 (1), 134.
SAFETY
Basic Safety Question for iGEM 2013
1. List and describe all new or modified coding regions you will be using in your project. (If you use parts from the 2013 iGEM Distribution without modifying them, you do not need to list those parts.)
| Part number. | Where did you get the physical DNA for this part (which lab, synthesis company, etc) | What species does this part originally come from? | What is the Risk Group of the species? | What is the function of this part,in its parent species ? | |
| Ex | BBa_C0040 | Synthesized, Blue Heron | Acinetobacter baumannii | 2 | Confers tetracycline resistance |
| 1 | BBa_K1070000 | PCR, Dr.Baishan Fang's lab, Xiamen University | Registry of Standard Biological Parts | 1 | The promoter induced by arabinose |
| 2 | BBa_K1070001 | PCR, Dr.Baishan Fang's lab, Xiamen University | Registry of Standard Biological Parts | 1 | The promoter induced by arabinose |
| 3 | BBa_K1070002 | PCR, Dr.Baishan Fang's lab, Xiamen University | Registry of Standard Biological Parts | 1 | The promoter induced by arabinose |
| 4 | BBa_K1070003 | PCR, Dr.Baishan Fang's lab, Xiamen University | Registry of Standard Biological Parts | 1 | The promoter induced by arabinose |
2. Highest Risk Group Listed : 1
3. Please describe the chassis organism(s) you will be using for this project. If you will be using more than one chassis organism, provide information on each of them:
| Species | Strain no/name | Risk Group | Risk group source link | Disease risk to humans? If so, which disease? | |
| Ex | E.coli(K 12) | NEB 10 Beta | 1 | www.absa.org/riskgroups/bacteria search.php?genus=&species=coli | Yes. May cause irritation to skin, eyes, and respiratory tract, may affect kidneys. |
| 1 | E.coli(K 12) | DH5α | 1 | http://www.absa.org/riskgroups/bacteriasearch.php?genus=Escherichia | Yes. May cause irritation to skin, eyes, and respiratory tract, may affect kidneys. |
| 2 | E.coli(B) | BL21 | 1 | http://www.absa.org/riskgroups/bacteriasearch.php?genus=Escherichia | Yes. May cause irritation to skin, eyes, and respiratory tract, may affect kidneys. |
4. Do the biological materials used in your lab work pose any of the following risks? Please describe.
a. Risks to the safety and health of team members or others working in the lab?
No. According to the Final Risk Assessment of the U.S. Environmental Protection Agency (EPA) (1997), Escherichia coli K-12 are not likely to pose any risk to human or animal health, to plants or to other microorganisms. In addition,We will use gloves and eye protected glasses during the experiments.
b. Risks to the safety and health of the general public , if released by design or by accident?
No. Escherichia coli K-12 are not likely to pose any risk to human or animal health, to plants or to other microorganisms. Although strains we use in our lab work have antibiotic genes, but they can still be killed by other common antibiotics.We only use E. coli and we will have disinfection after the experiment.
c. Risks to the environment, if released by design or by accident?
No. Since Escherichia coli K-12 are ubiquitous in the environment, they won't endanger any regular species in the environment.We only use E. coli and we will have disinfection after the experiment.
d. Risks to security through malicious misuse by individuals, groups, or countries?
No. We only have the directional optimization of promoters and use GFP to characterize them. GFP won't do any harm to the safety and health of the public and the environment.
5. If your project moved from a small- scale lab study to become widely used as a commercial/industrial product, what new risks might arise? (Consider the different categories of risks that are listed in parts a - d of the previous question.) Also, what risks might arise if the knowledge you generate or the met hods you develop became widely available? (Note: This is meant to be a somewhat open - ended discussion question.)
Using site-specific mutagenesis of genes might bring in safety problems, so strict treatment of waste is required in commercial production. Besides, the methods we use are widely used in the lab, and they won't cause any disadvantages if used properly.
6. Does your project include any design features to address safety risks? (For example: kill switches, auxotrophic chassis, etc.) Note that including such features is not mandatory to participate in iGEM, but many groups choose to include them.
No.
7. What safety training have you received (or plan to receive in the future)? Provide a brief description, and a link to your institution’s safety training requirements, if available.
Yes,every member of our laboratory received biosafety training including standard laboratory operation and WHO Biosafety Manual training before the experiments. All members learnt about the "C3H3"which is well known in the college of chemistry and chemical engineering. http://210.34.15.138/rule/10384_2_c_18.pdf
8. Under what biosafety provisions will / do you work?
a. Please provide a link to your institution biosafety guidelines .
http://210.34.15.138/rule/10384_2_c_18.pdf
b. Does your institution have an Institutional Biosafety Committee, or an equivalent group? If yes, have you discussed your project with them? Describe any concerns they raised with your project, and any changes you made to your project plan based on their review.
No, we haven't discussed our project with an Institutional Biosafety Committee, but we discussed the safety problem with our instructors and confirmed that this project is safe enough if experimental rules are strictly observed.
c. Does your country have national biosafety regulations or guidelines? If so, please provide a link to these regulations or guidelines if possible.
http://english.biosafety.gov.cn/Center/200511/t20051101_13462.htm
d. According to the WHO Biosafety Manual, what is the BioSafety Level rating of your lab? (Check the summary table on page 3, and the fuller description that starts on page 9.) If your lab does not fit neatly into category 1, 2, 3, or 4, please describe its safety features [see 2013.igem.org/Safety for help].
Level 1.
e. What is the Risk Group of your chassis organism(s), as you stated in question 1? If it does not match the BSL rating of your laboratory, please explain what additional safety measures you are taking.
Risk group 1.
Faculty Advisor Name:
Fang Baishan
Faculty Advisor Signature:
iGEM Software Division Coding Practices Questionnaire
1. Are you using the iGEM Software repository at github.com/igemsof tware? If you have instead stored your code elsewhere, please explain where and why you have put it there. If your code is not in the iGEM repository, are you using any version control system such as Git, CVS, or SVN?
E' Note is a web software using SVN and Brick Worker is biobrick evaluation software which will use the iGEM Software repository at github.com/igemsoftware.
2.User Data
a. Does your software store any private data supplied by the user? (For example: the user's name and email address, passwords, DNA sequences, circuit designs, etc.) If yes, please describe what kind of data is stored. If no, skip the rest of this question and move on to question 3.
Yes, E' Note stores users' name, email address, passwords and experiment data that they submit. And Brick Worker doesn't store any private data.
b. What is the URL or IP address where the user's private data is stored? Where is the physical computer or hard drive that contains the user's private data?
E' Note's URL is http://sae.sina.com.cn/ that store the user's private data. It is the SinaAppEngine which is a Cloud platform made by sina corporation that contains the user's private data.
c. Please describe any encryption, password protection, etc. that you use to protect the user's data. (It is not mandatory to have such protections, but if you do, describe them.)
Because we rent the SinaAppEngine to make E' Note's application active, the user's private data is whole stored in it's Cloud storage which will be protected by a group of Safety professionals.
3. Does your software include any other security features? Please describe them here.
No, E' Note is in virtue of SinaAppEngine to make the most powerful protection and Brick Worker is to evaluate and analyze the biobrick sequence and dose not include any storage of the user's data.
4. Does your software let the user create a design by choosing parts/genes from a list/database, such as the Registry? If so, which lists/databases are included? Is there any restriction on which parts/ genes the user can choose?
Yes. For Brick Worker, in the primer design part, we allow the user to choose transcription factors and add transcription factor binding sites (TFBS) to the primer sequence, and the TFBS data is derived from RegulonDB.
5. Does your software allow users to write new data into any public lists or databases? If so, do you check the new data for errors before allowing it to be written?
No.
6. Does your software include any other features that encourage th e user to create safe designs? Please describe them here.
Yes. Our Brick Worker help user to locate the essential part of promoter and RBS so the user can design mutation in these sites to improve the quality of the biobricks and in the protein coding sequence optimization part our software automatically optimize the protein coding sequence to a higher expression level.
OUTREACH
This year XMU_Software and XMU_China 2013 worked together to hold a series of programs of Human Practice. The Human Practice aims to further promote synthetic biology and popularize synthetic biology safety education. It also devotes to the promotion of International Genetically Engineered Machine Competition and academic exchanges with other domestic competitors, as well as propagandizing the two iGEM teams from XIAMEN University, XMU_Software, XMU_China 2013 and their respective projects.
Exchange of experience among domestic iGEM teams
      April 5, 2013, we had great honor to welcome iGEM team leader, ZhangHaoqian, a student from Peking University, to come to XIAMEN University in purpose of strengthening communications of experience between our two universities. During this time, Mr Zhang took part in discussions and brainstorming between our two teams, XMU_ Software and XMU_ China2013,and commented on the realization,security,advantages and disadvantages of ideas that put forward by our students. After that, Mr Zhang simply introduced previous projects as well as encouraged us all to make efforts for this year's iGEM projects, which benefited us tremendously.


              Student ZhangHaoqian introduces Peking University's iGEM                            ZhangHaoqian and iGEM team of Xiamen University
      June24, 2013, NANJING University's iGEM team visited our university in hope of promoting communications between us. Firstly, we showed them around natural attractions in our university. Then, we two schools’ iGEM teams gathered together for a meeting to exchange experience at night, in which the iGEM team leader from NANJING University firstly gave a brief introduction of their projects and then we also presented this year's projects, cell oscillation circuit and optimizing the orientation of the promoter. At the same time, we had intense discussions on the feasibility and safety of these projects. Finally, we extended our great longing and blessings for contests in the future for each other in the hope of making achievements in this year's contest.


      Friendly communication between Nanjing University and ours in the iGEM        Student from Nanjing University introduces their iGEM program


            Student from Xiamen University introduces their iGEM program                   iGEM teams from Nanjing University and Xiamen University
Lectures About Synthetic Biology
      During the summer camping, we organized a variety of Human Practice activities along with FuJian Chemical Society. In the evening of July16, 2013, we gave a series of lectures concerning about synthetic biology and iGEM for all the summer camp seniors from FuJian Province. These lectures mainly talked about what is synthetic biology, introduction of iGEM and its development in XiaMen University, together with seniors’ projects. We first tried our best to illustrate what is iGEM in every aspects, from basic ideas of synthetic biology, operation, development process to project classification, web pages and security. Then, we welcomed all the iGEM teams of XiaMen University to talk in detail about our long course of struggle for iGEM. After that, we gave a simply introduction about seniors’ part in iGEM which were closely concerned by high school students.
      During the lectures, we also made questionnaires about synthetic biology and iGEM as well as organized the mutual interlocution part, so that high school students could be in the joy of learning more knowledge.


                  Presentation about lectures of synthetic biology                           Member ZhangYijuan introduces synthetic biology


                         Enthusiastic interaction among seniors                                              Xiamen University's iGEM team members and seniors
Themed Garden Party of Synthetic Biology
      In order to publicize synthetic biology and iGEM, we held a themed garden party of synthetic biology in Guihua Mountain Building in our campus on July 17th 2013, after the cathedra. The party started at 2:00 pm and lasted for 3.5 hours, oriented to the high schools form summer camp of Fujian Province.
      Themed show of synthetic biology came first. We introduced systematically basic content of synthetic biology and its security issues to our high school audience, as well as ABCs of iGEM. Synthetic biology knowledge, safety and security principles, rules and regulations for operation were our chief presentation with the help of multimedia.
      Then games took their turn. We had lantern riddles, handing some synthetic biology riddles witch leads to proper nouns so as to help remembering, on lanterns. We had DNA difference spotting, putting two DNAs of tiny differences together as a challenge for insight and memory. Noticeably and proudly we create a new game in the foundation of Fold-it, a protein folding game. Scores will be marked on the basis of qualities of protein models.(link: http://verycd.gdajie.com/topics/2817908/) Last but not least, we alter and redesigned Sanguosha into a new card game: Synthetic-biology-sha. It was a combination of theories of experiment operation and bacteria, as a joyful way for our campers to learn more about synthetic biology.
      Except for that, our specially designed poster series were a great attraction which received high opinions from all folks.


          Seniors from summer camp take part in the game of lantern riddles                                        Game of protein folding


         Students are playing game of protein folding with intense enthusiasm                    Game scene in finding differences among DNA


                    Members of iGEM introduce Synthetic-biology-sha                                      Members of Xiamen University's iGEM team
Doodles of Synthetic Biology
       Furong Tunnel, one of the Xiamen University's main tourist attractions, visited by more than 1,000 people per day statistically, witnessed the doodles of iGEMers form Xiamen University. The doodle mainly includes the combination between experiment and assembly language, stressing the core of iGEM software team: assembly language for the software, experiment for the verification.

                                    iGEM paintings in Fu-Rong tunnel
      Our Human Practice was an acceleration for popularization of synthetic biology, iGEM and entries of Xiamen University for XMU_Software this year, an education of its safety and security for further step; a platform for academic exchange with other domestic iGEM teams. It was indeed a success as, if no more than, expected.
NOTEBOOK
TOOLS
01



XMind
PowerPoint
Robert's Rules of Order
XMind
The software was used for record our brainstorm and make new ideas. It is free! And it has so many models for new users to build up their own models.
PowerPoint
That Powerpoint is used for reporting our progress and giving presentations. Thus we practice it and study how to tell a funny story.
Robert's Rules of Order
This book guided us on how to make a meeting effective. And it really worth the hornor.
TOOLS
JOURNAL
02

MARCH
2013.3
2013.3.30
Chen:Experimental workspace that can automaticly decide whether it is a personal account or business account from the networking and the workspace afford download link of those relevant softwares.Can we get those softwares embedded? How to help our costumers find out the software they want quickly?(keyword search?) It is also important to processing data,generate lab reports and logs in the workspace.
Future plan:adding softwares in the software library and optimizing the software's functions and landscaping interface. Besides,we want to afford real time data while monitoring on different platforms.And how about sync to the cloud?
Likai Qiu:easy express. The purpose of design:economize time for researchers, and then they just need to stay on creating new ideas and putting possible idea into practice. Besides, unified expressions get researchers understand other's work easily.Its system's function:drawing,display sequence,dynamic display structure of the plasmid and parts of the experiments,shows the meaning of each graph and table,guiding and help.Question:can we really dynamic display the data? If we standardization the presentation, the software may be reject by other teams for its boredom standardization different kinds of efforts may be impossible.This software told you that you don't need to learn too MAYA and other magic softwares, it can include their funcitions, will it be possible?(perhaps impossible for now) By attach database to the software and afford individuation information search for our users.
Chuyue Wang:document management and real time monitoring that can help on scientific literature management and take notes and share your ideas.Automatic detection and data recording of streamlined repetition, design shortcut keys to improved operational efficiency. Using graph visualization to understand the data easily.
Yijuan Zhang:directional optimization of BioBricks.
Purpose:input enviromental parameters including special loci of genes, the expected expression level and chooses candidate BioBricks. optimize and find out the best sequence under given conditions and setting number of evolutionary (n) chooses the BioBricks, and the systerm will guide the user until the given conditions were met, variation to generates new sequences. Copy,retain or reject BioBricks according to the matched-degree, this step is parallel to natural selection. Circulate
Stop after circulate n times, output the best sequence. The problems are how to make sure that the genotype equals to its phenotype? How to define the f(x) that we want? And now we need to read some books about genetic algorithm to get the way.
Summary:Now we have two ideas,build a platform for researchers focusing on synthetic biology (aim to increase their efficency). Already several teams work on it, so it might be a rat race .
Solve synthetic biology problems with GA. We need to supplement the relevant basic knowledge of GA.So,studying CLOTHO developed by Berkeley and other universities and try to find out some new ideas and make a research to find out what front-line researchers need.We also need to study relevant softwares and GA,protein quaternary structure and knowledge about directional optimization, read more books and literature to make sure the possibility of the idea. For GA is too difficult for some of us, we still need to get come up with some new ideas.
OK, for this much tasks, work in groups and share new ideas~!
Meeting Record
3.30
APRIL
2013.4
2013.4.7
Zhang Yijuan had learn GA & the 《Alteredpeptideligands》,its diameters is in coden,but we need to improve BioBricks,so we need to change it to suit us and increase efficiency.Also ,we read news about Calgary and NewCastle ,SUSTC(calculate terminator efficiency by a score systerm).For coden is linked up with fitness,which influence the secondary structure of protein. For example, the changing of A-tail, stem loop and T-tail, we can also find it in the project of SUSTC.Besides,she also read papers on how to calculate f(x) & take notes when learn new things. SUSTC' fault:just one function, it may not catch everybody' eyes . She plan to change promoter so as to confirm the system efficiency.Huang Jianxing schedule to learn neural network ,after that,he will change the source code to be used on today's system.Huangxin and Huangjian now know just a little about matlab but they think the code can be change easily. Wang Chuyue learn matlab as well.Tangchun read some books about system efficiency and download papers to share on the group.From N-terminal domain rule, he notice when choose parameters is a key point for our project.
Next week's task had been written on the wall. And we suggest everybody upload and try to explain by powerpoint.
2013.4.14
Specific our goals:for most of us aren't familiar with GA, we misunderstand Yijuan's idea at the beginning. we determined to find the best fit BioBrick through genetic recombination and genovariation inside the BioBricks, and we evaluating them by our algorithm.Compare with other team' work:EbolGENTS(2008,Calgary) and Ebolutionary Algorithm(2008,Newcastle University). both of them had used GA, however ,they construct a system with given BioBricks just like build blocks, after which they find the best system in those "structures". as for our group, we focus on genetic recombination and genovariation inside the BioBricks to make new ones, and search for the best with given conditions.
Reduce the degree of difficulty:it may be a huge project to ensure phenotype and fitness result from mutations of base sequence in the whole BioBricks.so we decided to narrow down and aim at optimize decision of particular bases, promoter, terminator and RBS locus.Yijuan had mentioned that SUSTC had done research on computational efficiency of the terminator's gene sequence.
Core problem:the core problems of our idea contain two parts:How to ensure phenotype by genotype? How to find the f(x) meet our goal? Fitness function:three ways had been chosen to slove it: first, search for relevant literatures; read the project offered by Professor Fang named " microorganism's choice of the optimal conditions"; if possible, our wet lab should give us a hand on affording real time data and experimental simulation to ensure f(x).
Current task:check the project of iGEM "calculate terminator’s efficiency by sequence of the terminators" ;all of us need to take a look on SUSTC' work and discuss the next time.Learning Matlab and GA!
Annotations:we had set sail now!All of us need to learn SUSTC's work on ensuring genotype by phenotype and discuss the next time.We are badly in need of professor's help!
Try to find more idea! stay hungry!
2013.4.21
This week we had summary the work we had done before and nowadays we need to fix it.GA as a algorithm can work on several biological questions. And several new ways and improvement had been found out to slove the problem. Take an example, GASCO is a new algorithm that improve the answer of GA.
We start to plan the human practice. That may be of great importance. We had distribute the job and explicit the goal so as to lifting efficiency.
Neural network can be used in it.
2013.4.28
Zhang yijuan:after reading several papers and get down to SBOL. She got some new ideas on our program.
Han tao & Huang Jiang & Huang Xin: they had success got a software that can simulate the situation. However, the software remain some problem to be slove. We had better make a new one, do not use this one, for we can use Matlab.That will be a good idea.
The platform and relevant work went on well.
Meeting Record
4.7
4.14
4.21
4.28
MAY
2013.5
2013.5.5
Most of us take a rest after long hours.
2013.5.12
From this week,we will be divided into two groups, one named design group and another one Matlab group. For the first group, they need to finish most of work about design. Our logo,clothes and slogan will be their works. Besides,the platform and the wiki to show our achievement require them to try their best to do it. For the second group, they must continue working on the software and try to improve existing algorithm.
We had got some ideas on human practice and the next step was to check if they can be carried.
2013.5.19
Design group:the wiki has been designed and we are now considering combining wet lab's work with us. Han Chen had do a lot work in wet lab and found out what problems they met. We will finish the wiki in the next few days and afford some tools for users.
Matlab group:already written part of our software and test it. We test it and find that its interface is not friend and difficult for user to master it for the first sight. And we will call for help from the design group. Also,for the semi finals is nearby. We need to take a break.
2013.5.26
Huang Jiang &Huang Xin: They learn C++ and try to make a program . They learn different kinds of arithmetic and compare them to find what we need.GASCO and some other algorithm are inconsideration.
Zhang Yijuan:find softwares that can help us to slove those problems.Besides, she also focus on SBOL and find out the way to slove some easy problems we put forward before.
Zhang Shiqi & Wang Chuyue: already use GA to make a program and test it. But we think they need to make more progress so as to up to standard.
Qiu Likai & Chen Yuezeng:the software had been finished and they are design the wiki and writing codons. That might be finished in two months.
Huang Jianxing: read and find some papers about Codon usage bias. He should have been better.
NCBI is of great importance if we write a software ourselves. And we need to know how to make use of the data in NCBI.
Meeting Record
5.5
5.12
5.19
5.26
JUNE
2013.6
2013.6.2
Lin shen:codon bias database is important for our works. We need to use Latin to search for the right sequence for the organisms. NCBI had been downloaded and can be imported. PFD(protein folding kinetics), nowdays, 'rotational inertia" as a parameter that affect its folding speed.
Chen han:everything goes well. He start to prepare for the human practice.
Zhangyijuan:already explain the meaning of RBS. And figure out the way that how to evaluate it. That idea had been written and be used in our software.
Hon Yuming: draw something and had got the idea of doing so. The logo design is at the handle. And the logo design influence the next step and we shall finish it ASA possible
2013.6.9
Exam week. So call off.
2013.6.16
Exam week. So call off.
2013.6.23
Zhang yijuan had distinguish general transcription factors, recognition of inductivity transcription. Improving by similar softwares.Then she will import the database and check them one by one.So we need to know how to call PWM from the database.On sigma factor, statistic PSFM model by using neural network will be a good way. We know that best length require a certain distance.For not ready-made matrix,we had to written one.She will read papers about RBS following.Huang jianxing & Huang jiang & Lin shen & Han tao had read to know the basic knowledge.Cheng han find papers about sigma 70 & do some work on how to build an experience platform that suits users’s need. He improve and redo the platform to make a new model.
Team member Yijuan Zhang reported our software project (mainly about evaluating of promoters) to Mrs. Wu , the advisor of wet lab team . We received several suggestions from Mrs. Wu . To enhance the accuracy of our software work, we are supposed to pay attention to the false positives raised during recognition of promoters. And to further improve our promoter-evaluation program, Mrs. Wu suggested that we do some research on some special promoters besides Escherichia coli promoters. On the whole Wu applauded progress we had achieved and encouraged our software team to do our wonderful jobs as always.
Summer camp,garden party,cathedra of synthetic biology, Card game, Fold it and lantern riddles will help us to introduce iGEM to people.Financial reimbursement including registration fee. Several matters need attention: like travel expense,laboratory items,office supplies,car fare. Printing and copying cost should label amount of money.Qiu likai & Chen yuezhen made the concept map and interface about our software.Huang xin make use of the software that we written before. Then he will look for more papers about sigma 70.
2013.6.30
Arithmetic software team on evaluating and optimizing biobircks held a meeting and report our progress and current problems to former iGEMer Mo Youbin. Mo listened to our program with great concern and appreciation and taught one of our team members to us artificial neural net work to build weight position matrix for sigma motifs. We discussed the difficulties in front of us, which is mainly about the refinement of our algorithm and how to store and use the data from database RegulonDB. We decide to focus on improving the algorithm these a few days. In addition, Mo suggested that we start to present our slides in English and make a summary of our theories so as to post them on our wiki.
Easy Note is the software based on web, which is designed to convenient biosynthetic researchers to do some experimental record and query information. After the registration and login, users can do some basic records, related database and background program and so on.
Because the interface is too monotonous, and the function is too simple, we plan to design a succinctly and naturally interface and add query information, timing function, set, import and export document in the next month.
Meeting Record
6.2
6.9
6.16
6.23
6.30
JULY
2013.7
2013.7.7
Han designed small icon so that the interface will be more beautiful. Small icon included add, delete, document, tool, set, text, picture and so on.
Qiu had successfully realized login in and registry database. At the same time, he coded the data board. Chen had written code for the interface including add, delete, document, tool function. But, it is still not enough perfect because of various reason. What to do next will be discussed at tonight’s meeting.
Huang Xin learned that we can deal with the data from database using matlab.
Han Tao refined our algorithm to evaluate promoter strength on java and try to run some existing software designed to improve protein coding sequences.
Zhang Yijuan started to learn theories on RBS and its prediction. By looking up to literatures, we find several factors which can influence the RBS strength including SD sequence , spacer length, start codon and secondary structure. And we come up with algorithms similar to the algorithms used in the promoter part which take 3 factors, namely, SD sequence, spacer length, start codon into account. And we’ll study the influence of RBS structure on its strength next week.
Rao Wenjun has built a database in Access containing all the transcription factors’ binding sites and we are now discussing how to use this database in our software.
2013.7.14
To improve the login interface and registry interface, Chen designed a new style’s interface which had divided into four section and had used irregular design. When clicking one section, this section enlarged so that the interaction effect was better. Qiu made the great efford to improve the date base. He had finished the add and delete function. Because the date base is difficulty, he spent a lot of time for it so he stayed up late sometimes. Next week, we will improve the function and interface.
Zhang figured out the algorithm to calculate the free binding energy between RBS mRNA and 16s rRNA and HanTao started to write code for it at once. Now we have a simple program for calculating the free binding energy. So far we have worked out two algorithms to evaluate RBS strength.
2013.7.21
ZhangYijuan put forward 2 methods to evaluate RBS strength. The first method is to use the PWM(position weight matrix) of SD sequence and the spacer length between SD sequence and start codon to calculate the relative strength. The second is to calculate the free binding energy between the RBS mRNA and the 16s rRNA in the ribosome.
HanTao,HuangXin and RaoWenjun is working together on program code.
Qiu had finished the record module that we can add text, photo, table and so on. Because the record interface isn’t too concise, we should improve in the next week. At the same time, the import and export function had accomplished. In the process of programming, we met with many difficulties. For completing the task better in the next stage, Chen studied the jqery. Next week, we will continue to work hard.
2013.7.28
HanTao began to deal with the GUI of the promoter evaluation part of our project.
RaoWenjun decide to inlay the biobrick sequence data in our software as txt file.
ZhangYijuan had put forward the problem of how to determine the weight of motif similarity and spacer length score in the strength evaluation algorithm .After consulting with QiuRuosang, Zhang and QiuRuosang began to design experiments for the data needed to modify as well as validate our algorithm to evaluate both promoter and RBS strength.
After the end of the third semester in this week, we invest a lot of time for the software production. Qiu had finished and modified the function of adding photograph, table, step and the text. The difficulties in the database exceed we imagine. With increasing a lot of novel effect, login interface had been modified by Chen. Next week, we will tidy up the code for easier to read and improve the existing functionality.s
Meeting Record
7.7
7.14
7.21
7.28
AUGUST
2013.8
Chen:Experimental workspace that can automaticly decide whether it is a personal account or business account from the networking and the workspace afford download link of those relevant softwares.Can we get those softwares embedded? How to help our costumers find out the software they want quickly?(keyword search?) It is also important to processing data,generate lab reports and logs in the workspace.
Meeting Record
8.1
8.8
8.15
8.22
Lab Record
3.16
3.19
3.23
3.27
3.28
SEPTEMBER
2013.9
Chen:Experimental workspace that can automaticly decide whether it is a personal account or business account from the networking and the workspace afford download link of those relevant softwares.Can we get those softwares embedded? How to help our costumers find out the software they want quickly?(keyword search?) It is also important to processing data,generate lab reports and logs in the workspace.
Meeting Record
3.7
3.15
3.22
3.30
Lab Record
3.16
3.19
3.23
3.27
3.28
OCTOBER
2013.10
Chen:Experimental workspace that can automaticly decide whether it is a personal account or business account from the networking and the workspace afford download link of those relevant softwares.Can we get those softwares embedded? How to help our costumers find out the software they want quickly?(keyword search?) It is also important to processing data,generate lab reports and logs in the workspace.
Meeting Record
3.7
3.15
3.22
3.30
Lab Record
3.16
3.19
3.23
3.27
3.28