 "
"
Team:KU Leuven/Project/Oscillator/wetlab
From 2013.igem.org



Secret garden
Congratulations! You've found our secret garden! Follow the instructions below and win a great prize at the World jamboree!
- A video shows that two of our team members are having great fun at our favourite company. Do you know the name of the second member that appears in the video?
- For one of our models we had to do very extensive computations. To prevent our own computers from overheating and to keep the temperature in our iGEM room at a normal level, we used a supercomputer. Which centre maintains this supercomputer? (Dutch abbreviation)
- We organised a symposium with a debate, some seminars and 2 iGEM project presentations. An iGEM team came all the way from the Netherlands to present their project. What is the name of their city?
Now put all of these in this URL:https://2013.igem.org/Team:KU_Leuven/(firstname)(abbreviation)(city), (loose the brackets and put everything in lowercase) and follow the very last instruction to get your special jamboree prize!


The Sticker System
Integrating The Oscillator To The Wetlab
A well-known problem associated with the use of insect repellents is habituation. After a while insects will get used to the smell of the repellent and they won’t react anymore. Therefore we decided to implement an oscillator in our project for the production of (E)-β-farnesene (EBF) and methyl salicylate. In this part we will explain the genetics of our oscillator. For more information on the theoretical system and our in silico design, we refer to our design page and oscillator modelling page. We propose an implementation of the oscillator design and during the summer we focused on creating specifically a feed forward loop incorporating a quorum sensing molecule, since this is the workhorse of the oscillator.
How the oscillator works
We will shortly describe the functionality of the model as this is necessary to understand the resulting genetic network. Our proposed model is a network consisting of different transcription factors (TF), as is displayed in the figure. When there is a high level of TF A, this induces the production of both TF X and TF C, of which the latter will repress TF A and induce TF B. The production of TF C will not stop, even after TF A has disappeared below its threshold, since there is still TF X present that induces TF C. This effect creates an extended repression of TF A, in order to make sure the level of TF A drops sufficiently, so there is no production of TF X and TF C while the other halve of the system is active. This extended period in which TF C is present, also induces the production of TF B and the same story can be told for this halve. This means there are sequential peaks in the different components, which means this is an oscillating system.
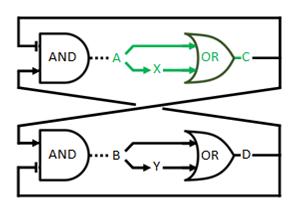
Figure 1 ǀ The oscillator model
From abstract network to biological implementation
As mentioned above we chose to use two quorum sensing molecules and four other transcription factors. A difficult step is the implementation of all the required interactions between those transcription factors. In our proposition we accomplish this by simply using four inducing transcription factors (on top of the two quorum sensing molecules). This section discusses how these can be used to create the OR gates and the AND gates.
How to implement an OR gate
The most elegant solution would be to design a promoter that would perfectly exhibit the characteristics of an OR gate. In the wetlab we started a slightly altered design, in which the goal of a asymmetric time delay is still achieved. For a further elaboration on that option we refer you to the design page. We do propose a potential implementation for a genuine OR gate. This would consist of a gene that has two promoters in front of it. There would then be production when either one of the transcription factors is present and the increase when both are present would be reduced. In the case of C this looks as follows:

Figure 2 | The proposed implementation of the OR gate.
How to implement the AND gate
We have also worked on a proposal to implement an AND gate. We need a promoter that is activated by one transcription factor and inhibited by another. An extra complication is that the transcription factor that inhibits has to activate another promoter and most transcription factors only exhibit either inhibitory behavior or inducing behavior. We propose two different solutions to tackle these issues, of which the second appeals to us the most. Our first solution consists of using a second gene that is activated by the transcription factor that was meant to inhibit the AND gate. This second gene creates antisense DNA of the eventual gene en thereby blocks the production of the protein. In the case of A this looks as follows:

Figure 3 | The first proposal for the AND gate.
Our second solution takes notice of the fact that both transcription factors that play a role in the AND gate are activating transcription factors. We do need to turn the function of one of those around. The method we propose for this is literally turning the operator site for that transcription factor around. When that transcription factor is present it will bind to its operator site and attract polymerases. That polymerase will not be able to transcribe the gene, since it is oriented in the wrong direction and the polymerase will simply obstruct the binding of any other polymerase. In the case of A this looks as follows:

Figure 4 | The second proposal for the AND gate.
Let's put names on it
On the first part of the modeling page of the oscillator we showed (at least in silico) that there is only a low sensibility of the oscillating behavior to the exact choice of promoters and transcription factors. This study is far from complete, but we did not wait until the engineers were completely certain and put names on it. We have used several parts from the iGEM registry and different transcription factors with a widespread use. This results in the following scheme that exhibits the behavior as discussed in the design page and validated in the oscillator modelling page.

Figure 5 | The total implementation of the oscillator.
This would bring the total implementation to 7 constructs which is, at this stage in the evolution of synthetic biology still a tough task. This will become easily feasible when the artificial synthesis of DNA becomes more available and when the concept of a ‘minimal cell’ is attained. That is why we only focused on creating a feed forward loop.
Focusing on the feed forward loop
The genes
Since we want a colony-wide synchronization, TF A has to be a quorum-sensing molecule (as well as TF B). We chose for the autoinducer synthetase (BBa_C0076) that leads to the production of the quorum-sensing molecule 3OH,C14:1-HSL. Before this gene we placed a TetR repressible promoter (BBa_R0040), so the quorum-sensing molecule is only produced if there is no TetR present. Because this is a proof of concept, we want to be able to control the production of the quorum-sensing molecule. Therefore we placed the gene coding for TetR (BBa_P0340) behind a T7 promotor (BBa_I719005). When we put the construct in a Rosetta strain, we can control the activity of the T7 promotor. This strain has a chromosomal copy of the T7 RNA polymerase gene behind a lacUV5 promotor, which can be induced by IPTG. By adding IPTG, T7 RNA polymerase will be formed, which leads to the transcription of the TetR gene so no quorum-sensing molecules will be produced.
To be active, 3OH,C14:1-HSL has to form a complex with the CinR activator. This complex can bind the promotor BBa_R0078 and activate transcription. The gene for the CinR activator (BBa_C0077) is also placed under control of a TetR repressible promotor. So if no TetR is present, CinR and 3OH,C14:1-HSL are formed and they combine to activate the promotor BBa_C0078 which is placed before the gene coding for AraC (BBa_C0080; representing TF X) and before a gene for GFP (BBa_K082003; representing TF C). So the complex makes sure that TF X and TF C are formed. In the real model TF C would be the gene for EBF, but in our proof of concept model we use GFP, which allows us to easily detect if the system works.

Figure 6 | Genetic network of the oscillator.
Our control mechanism does not only consist of TetR, but we also inserted a gene in our model that codes for the autoinducer inactivation enzyme AiiA (BBa_C0060). This enzyme can hydrolyze the quorum-sensing molecule. The gene is also placed behind a T7 promotor. If IPTG is added, no quorum-sensing molecule and receptor are formed anymore and the remaining quorum-sensing molecules are degraded by AiiA.
AraC also leads to the transcription of the GFP-gene by binding to an AraC regulated promotor (BBa_R0080). This means that not only TF A leads to formation of TF C but TF X as well. We want to prove in the lab that if TF X also activates production of TF C, the cell will fluoresce for a longer period than if TF X is excluded from the network.
Results
As you can see on figure 2, the proof of concept of the oscillator exists of a lot of different genes. We started with creating TF C, which in our case exists of the GFP gene coupled to a promoter and a terminator. We ligated the GFP-reporter gene (BBa_K082003) to a double terminator (BBa_B0015). These form the new intermediate brick BBa_K1060006. On figure 3 you can see the insert and backbone after digestion of this new brick. Afterwards we focused on TF A, which consists of the genes coding for the auto-inducer synthetase and a receptor that binds the quorum-sensing molecule. We coupled the cinI gene which codes for the autoinducer synthetase (BBa_C0076) to the double terminator (BBa_B0015). These form the new intermediate brick BBa_K1060007.

Figure 7 ǀ Digestion of our new brick BBa_K1060006