 "
"
Team:Tokyo Tech/Project/Ninja State Switching
From 2013.igem.org
| (7 intermediate revisions not shown) | |||
| Line 9: | Line 9: | ||
[[Image:Titech2013_Ninja_State_Switching_2-1_1-2.jpg|350px|thumb|right|Fig. 2-1-2. Switching two states of <i>E. ninja</i>]] | [[Image:Titech2013_Ninja_State_Switching_2-1_1-2.jpg|350px|thumb|right|Fig. 2-1-2. Switching two states of <i>E. ninja</i>]] | ||
<p> | <p> | ||
| - | Through the human practice, we felt the importance of the explanation of the genetic programming in synthetic biology with interesting story. Thus we decided to make <i>E. coli</i> play | + | Through the human practice, we felt the importance of the explanation of the genetic programming in synthetic biology with interesting story. Thus we decided to make <i>E. coli</i> play "Ninja" story. In this story, there are three characters, so we prepared three types of <i>E. coli</i> which have different plasmids for each role: one is the hero, <i>E. ninja</i> and the others are <i>E. civilian</i> and <i>E. samurai</i> (Fig. 2-1-1). |
</p> | </p> | ||
<br> | <br> | ||
<p> | <p> | ||
| - | <i>E. ninja</i> changes between two states depending on whether there is <i>E. civilian</i> or <i>E. samurai</i> (Fig. 2-1-2). First, <i>E. civilian</i> is around <i>E. ninja</i>. <i>E. civilian</i> emits 3OC6HSL, a small intercellular communication molecule. When <i>E. ninja</i> detects 3OC6HSL, <i>E. ninja</i> switches into the | + | <i>E. ninja</i> changes between two states depending on whether there is <i>E. civilian</i> or <i>E. samurai</i> (Fig. 2-1-2). First, <i>E. civilian</i> is around <i>E. ninja</i>. <i>E. civilian</i> emits 3OC6HSL, a small intercellular communication molecule. When <i>E. ninja</i> detects 3OC6HSL, <i>E. ninja</i> switches into the "Mimic state" (Fig. 2-1-2 Step1). <i>E. ninja</i> maintains the Mimic state even after <i>E. civilian</i> goes away (Fig. 2-1-2 Step2). Then the <i>E. samurai</i> comes. <i>E. samurai</i> emits a different small intercellular communication molecule 3OC12HSL. When <i>E. ninja</i> detects 3OC12HSL, <i>E. ninja</i> switches into the "Attack state" (Fig. 2-1-2 Step3). <i>E. ninja</i> continues to maintain Attack state even after the <i>E. samurai</i> leaves (Fig. 2-1-2 Step4). Finally, <i>E. civilian</i> returns. <i>E. ninja</i> detects 3OC6HSL again, and switches back to the Mimic state (Fig. 2-1-2 Step5). |
</p><br> | </p><br> | ||
[[Image:Titech2013_Ninja_State_Switching_2-1_1-3.jpg|500px|thumb|right|Fig. 2-1-3. Ninja Circuit:Signal-dependent state change circuit with crosstalk circumvention]] | [[Image:Titech2013_Ninja_State_Switching_2-1_1-3.jpg|500px|thumb|right|Fig. 2-1-3. Ninja Circuit:Signal-dependent state change circuit with crosstalk circumvention]] | ||
<p> | <p> | ||
| - | To try to put this story into practice, we designed two genetic circuits, which we named | + | To try to put this story into practice, we designed two genetic circuits, which we named "Naive Circuit: Signal-dependent state change circuit" and "Ninja Circuit: Signal-dependent state change circuit with crosstalk circumvention" (Fig. 2-1-3). Both of the circuits have a toggle switch subcircuit and a toggle-repressor overexpression subcircuit induced by the intercellular communication molecules. In the case without crosstalk, each of the intercellular communication molecules, 3OC6HSL and 3OC12HSL, can switch the state of the toggle as reported by Collins group (H. Kobayashi et al., 2004). However, crosstalk between 3OC12HSL-LasR complex and <i>lux</i> promoter disrupts our scenario. |
</p><br> | </p><br> | ||
[[Image:Titech2013_Ninja_State_Switching_2-1_1-4.jpg|250px|thumb|right|Fig. 2-1-4. The result of crosstalk circumvention([http://parts.igem.org/Part:BBa_K1139110 BBa_K1139110])]] | [[Image:Titech2013_Ninja_State_Switching_2-1_1-4.jpg|250px|thumb|right|Fig. 2-1-4. The result of crosstalk circumvention([http://parts.igem.org/Part:BBa_K1139110 BBa_K1139110])]] | ||
| Line 29: | Line 29: | ||
<h1>2. Gene Circuit in Theory</h1><h2> | <h1>2. Gene Circuit in Theory</h1><h2> | ||
<p> | <p> | ||
| - | We designed two genetic circuits: | + | We designed two genetic circuits: "Naive Circuit: Signal-dependent state change circuit" and "Ninja Circuit: Signal-dependent state change circuit with crosstalk circumvention". |
</p> | </p> | ||
</h2> | </h2> | ||
| Line 35: | Line 35: | ||
[[Image:Titech2013_Ninja_State_Switching_2-1_2-1-1.jpg|500px|thumb|center|Fig. 2-1-6. Naive Circuit: Signal-dependent state change circuit]] | [[Image:Titech2013_Ninja_State_Switching_2-1_2-1-1.jpg|500px|thumb|center|Fig. 2-1-6. Naive Circuit: Signal-dependent state change circuit]] | ||
<p> | <p> | ||
| - | Originally, we designed the Naive Circuit: Signal-dependent state change circuit by combination with a bistable | + | Originally, we designed the Naive Circuit: Signal-dependent state change circuit by combination with a bistable "toggle switch"(T. Gardner et al., 2000)[1] and a toggle-repressor overexpression partial circuit induced by intercellular communication molecules (Fig. 2-1-6). Small molecules 3OC6HSL and 3OC12HSL can switch the state of the toggle as expressed in the following step-by-step descriptions. The toggle switch has two promoter-gene sets <i>lambda</i> promoter and <i>lac</i> promoter. <i>lambda</i> promoter and <i>lac</i> promoter are repressed by CI and LacI, and coding regions of these proteins CI and LacI are downstream of each promoter gene (Fig. 2-1-6). With the combination of the two subcircuits, our genetic circuit changes between two stable states by inducing intercellular molecules. Note that the explanation in below is in a situation without crosstalk problem which we address in the latter part of this page. |
</p><br> | </p><br> | ||
<div class="stepbox" id="step1"> | <div class="stepbox" id="step1"> | ||
| Line 93: | Line 93: | ||
<h3>3-1. Introduction</h3> | <h3>3-1. Introduction</h3> | ||
<h2> | <h2> | ||
| - | [[Image:Titech2013_Ninja_State_Switching_2-1_3-1.jpg|500px|thumb|center|Fig. 2-1-16. Summary of | + | [[Image:Titech2013_Ninja_State_Switching_2-1_3-1.jpg|500px|thumb|center|Fig. 2-1-16. Summary of "Crosstalk Circumvention Switch"]] |
<p> | <p> | ||
Our purpose is to check whether the <i>lux/tet</i> hybrid promoter would be repressed or not when 3OC12HSL-LasR complex and TetR are co-existed (Fig. 2-1-16). 3OC12HSL-LasR-dependent activation of <i>lux</i> promoter is known to be a problem in synthetic biology and we confirmed this crosstalk activation (Fig. 2-1-16). Then we compared the amount of crosstalk for <i>lux/tet</i> hybrid promoter in the presence or absence of the aTc, the TetR inhibitor. The binding between TetR protein and <i>tetO</i> sequence on DNA is known to be weakened by aTc. Tokyo Tech 2012 indeed showed that the GFP expression of the cells in which both of 3OC6HSL and aTc were added was higher than that of the cells in which only 3OC6HSL was added. Similarly, if the GFP expression of the cells we added both of 3OC12HSL and aTc was higher than that of the cells which we added only 3OC12HSL, it is proved that the crosstalk can be suppressed by TetR. | Our purpose is to check whether the <i>lux/tet</i> hybrid promoter would be repressed or not when 3OC12HSL-LasR complex and TetR are co-existed (Fig. 2-1-16). 3OC12HSL-LasR-dependent activation of <i>lux</i> promoter is known to be a problem in synthetic biology and we confirmed this crosstalk activation (Fig. 2-1-16). Then we compared the amount of crosstalk for <i>lux/tet</i> hybrid promoter in the presence or absence of the aTc, the TetR inhibitor. The binding between TetR protein and <i>tetO</i> sequence on DNA is known to be weakened by aTc. Tokyo Tech 2012 indeed showed that the GFP expression of the cells in which both of 3OC6HSL and aTc were added was higher than that of the cells in which only 3OC6HSL was added. Similarly, if the GFP expression of the cells we added both of 3OC12HSL and aTc was higher than that of the cells which we added only 3OC12HSL, it is proved that the crosstalk can be suppressed by TetR. | ||
| Line 100: | Line 100: | ||
<h3>3-2. Construction</h3> | <h3>3-2. Construction</h3> | ||
<h2> | <h2> | ||
| - | [[Image:Titech2013_CrosstalkCircumventionAssay_3-2_2.jpg|500px|thumb|center|Fig. 2-1-17. Gene circuits of | + | [[Image:Titech2013_CrosstalkCircumventionAssay_3-2_2.jpg|500px|thumb|center|Fig. 2-1-17. Gene circuits of "Crosstalk Circumvention Switch"]] |
<p> | <p> | ||
| - | We made a simple crosstalk circumvention system and named it | + | We made a simple crosstalk circumvention system and named it "Crosstalk Circumvention Switch" (Fig. 2-1-17). To construct the circuit shown in above, we ligated Pcon-RBS-<i>lasR</i>-TT ([http://parts.igem.org/Part:BBa_K553003 BBa_K553003]) and Plux/tet-RBS-<i>GFP</i>-TT ([http://parts.igem.org/Part:BBa_K934025 BBa_K934025]) as a reporter plasmid. We used Pcon-RBS-<i>luxR</i>-TT-Ptrc-RBS-<i>tetR</i>-TT as the regulator plasmid. |
</p> | </p> | ||
</h2> | </h2> | ||
| Line 132: | Line 132: | ||
<gallery widths="350px" heights="250px" style="margin-left:auto; margin-right:auto; text-align: center;"> | <gallery widths="350px" heights="250px" style="margin-left:auto; margin-right:auto; text-align: center;"> | ||
| - | Image: | + | Image:Titech2013_CrosstalkCircumventionAssay_3-2_5.jpg|<h2 style="font-weight: bold; font-size: 94%;">Fig. 2-1-19. Activation of our crosstalk circumvention system circuit</h2> |
| - | Image:Titech2013_Ninja_State_Switching_2-1_4-1.jpg.png|Fig. 2-1-20. The change of each protein concentration in switching from the Attack state to the Mimic state | + | Image:Titech2013_Ninja_State_Switching_2-1_4-1.jpg.png|<h2 style="font-weight: bold; font-size: 94%;">Fig. 2-1-20. The change of each protein concentration in switching <br>from the Attack state to the Mimic state</h2> |
</gallery> | </gallery> | ||
| + | |||
<p> | <p> | ||
During switching from the Attack state to the Mimic state, absence of TetR allows activation of <i>lux/tet</i> hybrid promoter. Protein TetR repressed by CI434, is indeed an important factor in the circuit. Under the Mimic state, TetR accumulates to plateau level. This presence of protein TetR is important to prevent crosstalk by 3OC12HSL-LasR complex. | During switching from the Attack state to the Mimic state, absence of TetR allows activation of <i>lux/tet</i> hybrid promoter. Protein TetR repressed by CI434, is indeed an important factor in the circuit. Under the Mimic state, TetR accumulates to plateau level. This presence of protein TetR is important to prevent crosstalk by 3OC12HSL-LasR complex. | ||
| Line 147: | Line 148: | ||
</p> | </p> | ||
<p> | <p> | ||
| - | We compared two circuits about the characteristics of changing from the LacI state to the CI state. The reason why we won’t compare them in the opposite way (changing from the CI state to the LacI state) is that crosstalk won’t occur in this direction. We analyzed the case that the initial state is with enough LacI, then it change to the state with enough CI at 300 min. by the increase of 3OC12HSL emitted from <i>E. samurai</i>. Fig. 2-1-22 shows the changing of LacI and CI. The solid line presents the case with crosstalk circumvention circuit, and the dotted line stands for the case without crosstalk circumvention circuit. When there is certain amount of 3OC12HSL production, LacI is produced in a certain amount in the toggle without crosstalk prevention circuit, and in contrary, the crosstalk would not be conspicuous. When <i>E. samurai</i> has gone at 1500 min., the concentration of LacI expressed by | + | We compared two circuits about the characteristics of changing from the LacI state to the CI state. The reason why we won’t compare them in the opposite way (changing from the CI state to the LacI state) is that crosstalk won’t occur in this direction. We analyzed the case that the initial state is with enough LacI, then it change to the state with enough CI at 300 min. by the increase of 3OC12HSL emitted from <i>E. samurai</i>. Fig. 2-1-22 shows the changing of LacI and CI. The solid line presents the case with crosstalk circumvention circuit, and the dotted line stands for the case without crosstalk circumvention circuit. When there is certain amount of 3OC12HSL production, LacI is produced in a certain amount in the toggle without crosstalk prevention circuit, and in contrary, the crosstalk would not be conspicuous. When <i>E. samurai</i> has gone at 1500 min., the concentration of LacI expressed by "Ninja circuit" is about 25 nM. In contrast, the concentration of LacI expressed by the original circuit is 725 nM. Regarding the convergence of LacI, we can conclude that LacI expressed by "Ninja circuit" will be converged faster than that of the original circuit does. |
</p> | </p> | ||
Latest revision as of 23:06, 28 October 2013

Ninja State Switching
Contents |
1. Introduction

 Fig. 2-1-1. Three characters in this story
Fig. 2-1-1. Three characters in this story
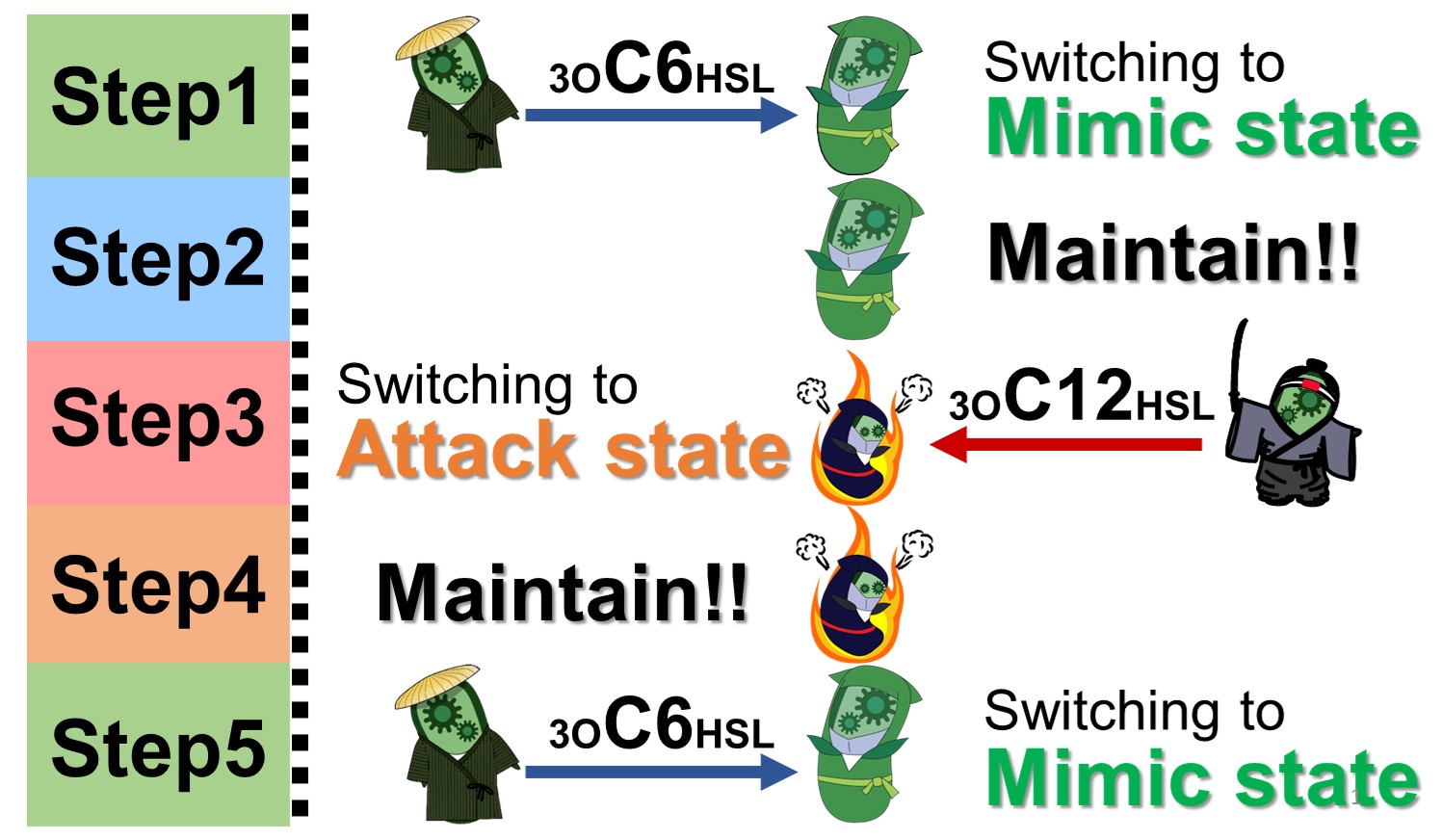 Fig. 2-1-2. Switching two states of E. ninja
Fig. 2-1-2. Switching two states of E. ninja


Through the human practice, we felt the importance of the explanation of the genetic programming in synthetic biology with interesting story. Thus we decided to make E. coli play "Ninja" story. In this story, there are three characters, so we prepared three types of E. coli which have different plasmids for each role: one is the hero, E. ninja and the others are E. civilian and E. samurai (Fig. 2-1-1).
E. ninja changes between two states depending on whether there is E. civilian or E. samurai (Fig. 2-1-2). First, E. civilian is around E. ninja. E. civilian emits 3OC6HSL, a small intercellular communication molecule. When E. ninja detects 3OC6HSL, E. ninja switches into the "Mimic state" (Fig. 2-1-2 Step1). E. ninja maintains the Mimic state even after E. civilian goes away (Fig. 2-1-2 Step2). Then the E. samurai comes. E. samurai emits a different small intercellular communication molecule 3OC12HSL. When E. ninja detects 3OC12HSL, E. ninja switches into the "Attack state" (Fig. 2-1-2 Step3). E. ninja continues to maintain Attack state even after the E. samurai leaves (Fig. 2-1-2 Step4). Finally, E. civilian returns. E. ninja detects 3OC6HSL again, and switches back to the Mimic state (Fig. 2-1-2 Step5).

To try to put this story into practice, we designed two genetic circuits, which we named "Naive Circuit: Signal-dependent state change circuit" and "Ninja Circuit: Signal-dependent state change circuit with crosstalk circumvention" (Fig. 2-1-3). Both of the circuits have a toggle switch subcircuit and a toggle-repressor overexpression subcircuit induced by the intercellular communication molecules. In the case without crosstalk, each of the intercellular communication molecules, 3OC6HSL and 3OC12HSL, can switch the state of the toggle as reported by Collins group (H. Kobayashi et al., 2004). However, crosstalk between 3OC12HSL-LasR complex and lux promoter disrupts our scenario.

In order to circumvent the crosstalk, we thus introduced two modifications. One is replacement of lux promoter for the overexpression with lux/tet hybrid promoter produced by Tokyo tech 2012 ([http://parts.igem.org/Part:BBa_K934024 BBa_K934024]). The other is addition of a repressor network containing cI434 and tetR. We confirmed crosstalk circumvention on the hybrid promoter experimentally (Fig. 2-1-4) and also showed the circumvention in the whole network by mathematical modeling (Fig. 2-1-5). The results correspond to the following scenario. When E. ninja receives 3OC6HSL, 3OC6HSL-LuxR complex activates lux/tet hybrid promoter, and E. ninja switches into LacI dominant Mimic state. On the other hand, when E. ninja receives 3OC12HSL, 3OC12HSL-LasR complex activates las promoter, and E. ninja switches into CI dominant Attack state (Fig. 2-1-3) because of crosstalk inhibition by tetR expressed in the LacI-dominant Mimic state.

2. Gene Circuit in Theory
We designed two genetic circuits: "Naive Circuit: Signal-dependent state change circuit" and "Ninja Circuit: Signal-dependent state change circuit with crosstalk circumvention".
2-1. Naive Circuit: Signal-dependent state change circuit
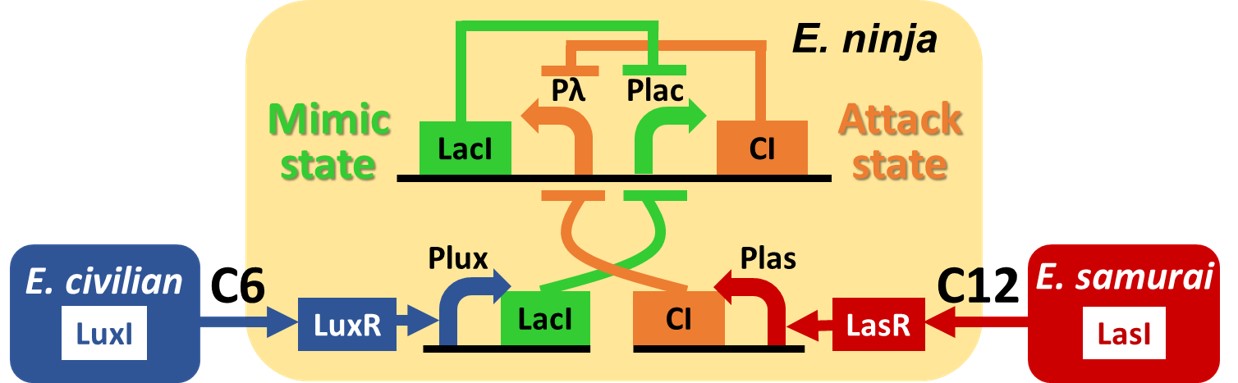 Fig. 2-1-6. Naive Circuit: Signal-dependent state change circuit
Fig. 2-1-6. Naive Circuit: Signal-dependent state change circuit

Originally, we designed the Naive Circuit: Signal-dependent state change circuit by combination with a bistable "toggle switch"(T. Gardner et al., 2000)[1] and a toggle-repressor overexpression partial circuit induced by intercellular communication molecules (Fig. 2-1-6). Small molecules 3OC6HSL and 3OC12HSL can switch the state of the toggle as expressed in the following step-by-step descriptions. The toggle switch has two promoter-gene sets lambda promoter and lac promoter. lambda promoter and lac promoter are repressed by CI and LacI, and coding regions of these proteins CI and LacI are downstream of each promoter gene (Fig. 2-1-6). With the combination of the two subcircuits, our genetic circuit changes between two stable states by inducing intercellular molecules. Note that the explanation in below is in a situation without crosstalk problem which we address in the latter part of this page.

Step1. When E. civilian emits 3OC6HSL and E. ninja receives it, the 3OC6HSL-LuxR complex activates lux promoter. LuxR is expressed constitutively in E. ninja, so 3OC6HSL combines LuxR when 3OC6HSL is received by E. ninja. Then LacI starts to be expressed because lux promoter is activated. And then LacI represses lac promoter. Finally, E. ninja switches into the Mimic state where LacI is expressed from lux promoter and lambda promoter on toggle switch (Fig. 2-1-7) (H. Kobayashi et al., 2004)[3].

Step2. When E. civilian leaves, lux promoter activation disappears because LuxR cannot form complex. However, LacI continues to be expressed from lambda promoter on toggle switch. Therefore E. ninja maintains the Mimic state (Fig. 2-1-8).

Step3. Next, E. samurai comes along. When E. samurai emits 3OC12HSL and E. ninja receives it, the 3OC12HSL-LasR complex activates las promoter. LasR is expressed constitutively in E. ninja, so 3OC12HSL combines LasR when 3OC12HSL is received by E. ninja. Then CI starts to be expressed because las promoter is activated and CI repress lambda promoter. Finally, E. ninja switches into the Attack state where CI is expressed from las promoter and lac promoter on toggle switch (Fig. 2-1-9).

Step4. When E. samurai leaves, las promoter activation disappears because LasR cannot form the complex. However, CI continues to be expressed from lac promoter on toggle switch. Therefore E. ninja maintains the Attack state (Fig. 2-1-10).

Step5. When E. civilian returns, E. ninja switches into the Mimic state again as same as step1. When E. civilian emits 3OC6HSL and E. ninja receives it, the 3OC6HSL-LuxR complex activates lux promoter. Then LacI starts to be expressed because lux promoter is activated by 3OC6HSL-LuxR complex. And then LacI represses lac promoter. Finally, E. ninja switches into the Mimic state where LacI is expressed from lux promoter and lambda promoter on toggle switch (Fig. 2-1-11).
2-2. The problem: crosstalk
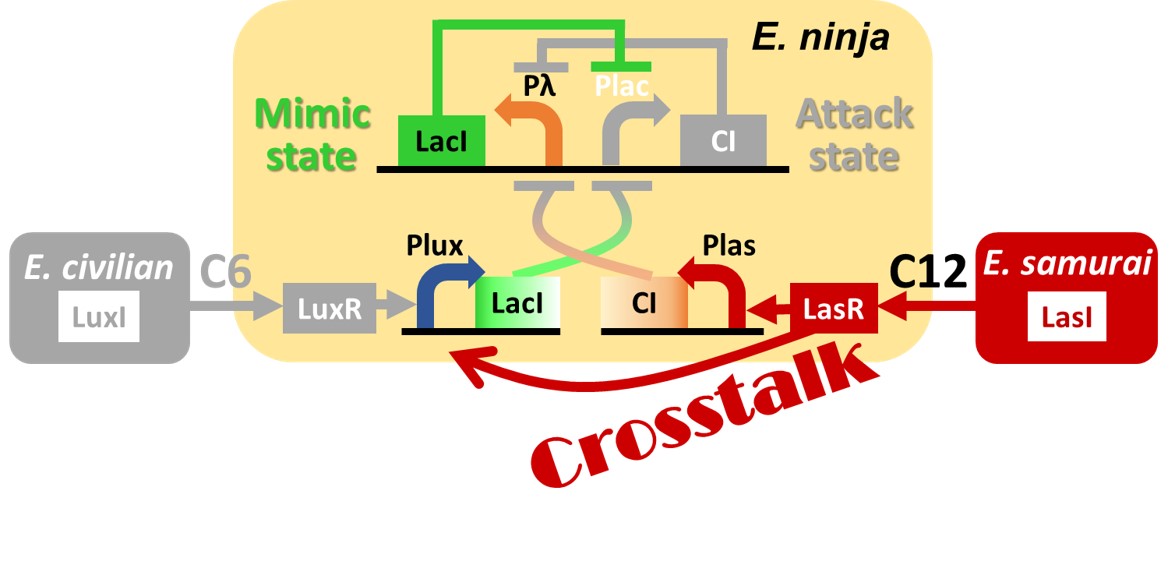 Fig. 2-1-12. This genetic circuit had a crosstalk problem
Fig. 2-1-12. This genetic circuit had a crosstalk problem

Though we described an ideal case in the above description, this genetic circuit had a crosstalk problem. In the case of this genetic circuit, 3OC12HSL-LasR complex, which turns E. ninja into the Attack state, activates not only las promoter but also lux promoter. The crosstalk of 3OC12HSL-LasR complex confuses E. ninja when E. samurai comes. When E. ninja received intercellular molecules 3OC12HSL from E. samurai, 3OC12HSL-LasR complex activates las promoter and also activates lux promoter (Fig. 2-1-12).

The crosstalk between the two intercellular molecules 3OC6HSL and 3OC12HSL has been reported in a scientific paper (Gray KM et al., 1994)[2]. We also quantified the crosstalk to confirm whether 3OC12HSL-LasR complex actually activates both las promoter and lux promoter (Fig. 2-1-12). We prepared two different plasmids: one has GFP regulated by las promoter and the other has GFP regulated by lux promoter. We introduced each plasmid into E. coli which has another plasmid to express LasR constitutively. Then, we added 3OC6HSL or 3OC12HSL to the culture. Results of this experiment are shown in Fig. 2-1-13. When we added 3OC6HSL, we could not confirmed adequate expression of GFP in both strains. Results show that little crosstalk exists between 3OC6HSL and LasR. On the other hand, when adding 3OC12HSL, we confirmed expression of GFP in both strains. This shows that 3OC12HSL-LasR complex activates not only las promoter but also lux promoter due to the crosstalk. (Crosstalk Confirmation Assay).
2-3. Ninja Circuit: Signal-dependent state change circuit
with crosstalk circumvention
We thought of two possible approaches to solve the crosstalk. One is protein/promoter engineering and the other is gene network engineering. We decided to choose gene network engineering to solve crosstalk problem.


We designed a new genetic circuit shown in Fig. 2-1-14. Our new gene circuit can circumvent crosstalk between the intercellular molecules 3OC6HSL and 3OC12HSL (Fig. 2-1-14). Two new genes cI434 and tetR are added to the original signal-dependent state change circuit. In addition, the lux promoter is changed to the lux/tet hybrid promoter, which is repressed by TetR. In the case when changing from the Mimic state to the Attack state under the presence of TetR, which is due to the absence of CI434, inhibits expression from the hybrid promoter. Without this inhibition, LasR protein, activated by 3OC12HSL from E. samurai, binding to LuxR-binding sequence of the hybrid promoter, would stimulate LacI expression from the promoter. This expression, in addition to the CI expression from the las promoter makes E. ninja confuse. On the other hand, using a new gene network, crosstalk is circumvented and E. ninja switches from the Mimic state into the Attack state normally. This is because lux/tet hybrid promoter is repressed by TetR (Fig. 2-1-15). Contrarily, in the Attack state, lux/tet hybrid promoter is not repressed due to the absence of TetR. This is because expression of TetR is repressed by CI434. So when E. civilian comes, lux/tet hybrid promoter is activated by 3OC6HSL-LuxR complex, then E. ninja switches into the Mimic state.
3. Crosstalk circumvention assay
3-1. Introduction
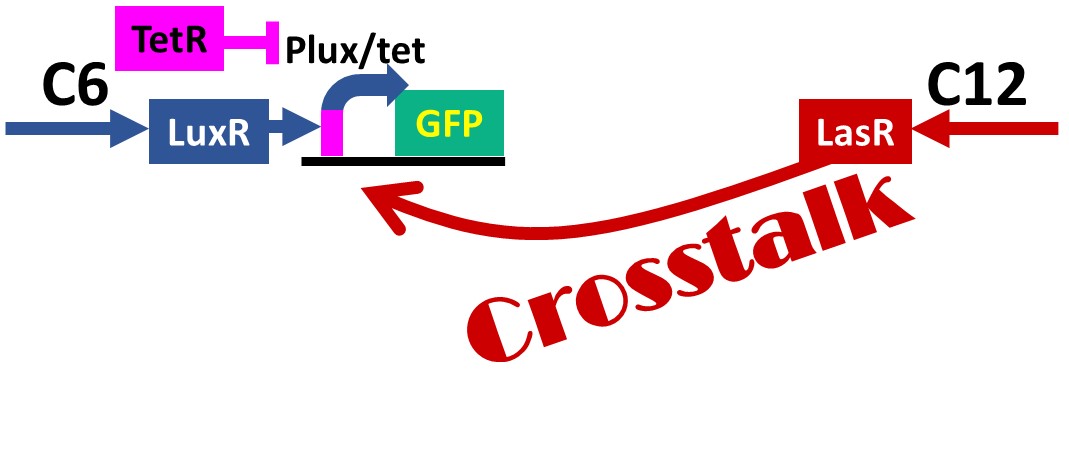 Fig. 2-1-16. Summary of "Crosstalk Circumvention Switch"
Fig. 2-1-16. Summary of "Crosstalk Circumvention Switch"

Our purpose is to check whether the lux/tet hybrid promoter would be repressed or not when 3OC12HSL-LasR complex and TetR are co-existed (Fig. 2-1-16). 3OC12HSL-LasR-dependent activation of lux promoter is known to be a problem in synthetic biology and we confirmed this crosstalk activation (Fig. 2-1-16). Then we compared the amount of crosstalk for lux/tet hybrid promoter in the presence or absence of the aTc, the TetR inhibitor. The binding between TetR protein and tetO sequence on DNA is known to be weakened by aTc. Tokyo Tech 2012 indeed showed that the GFP expression of the cells in which both of 3OC6HSL and aTc were added was higher than that of the cells in which only 3OC6HSL was added. Similarly, if the GFP expression of the cells we added both of 3OC12HSL and aTc was higher than that of the cells which we added only 3OC12HSL, it is proved that the crosstalk can be suppressed by TetR.
3-2. Construction
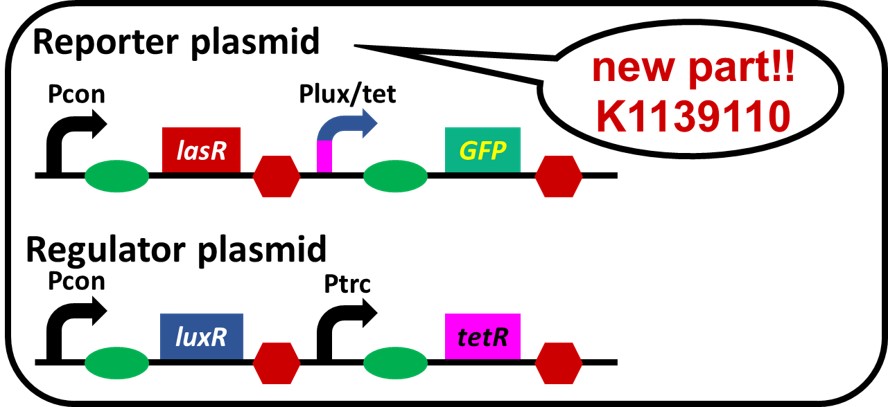 Fig. 2-1-17. Gene circuits of "Crosstalk Circumvention Switch"
Fig. 2-1-17. Gene circuits of "Crosstalk Circumvention Switch"

We made a simple crosstalk circumvention system and named it "Crosstalk Circumvention Switch" (Fig. 2-1-17). To construct the circuit shown in above, we ligated Pcon-RBS-lasR-TT ([http://parts.igem.org/Part:BBa_K553003 BBa_K553003]) and Plux/tet-RBS-GFP-TT ([http://parts.igem.org/Part:BBa_K934025 BBa_K934025]) as a reporter plasmid. We used Pcon-RBS-luxR-TT-Ptrc-RBS-tetR-TT as the regulator plasmid.
3-3. Results
In the graph below (Fig. 2-1-18), the level of GFP expression in cells where TetR is active is clearly lower than that of when TetR is inhibited. We confirmed this fact both in 3OC12HSL and 3OC6HSL. In short, the graph below shows that lux/tet hybrid promoter is repressed by TetR precisely. Furthermore, the graph below shows that there is a great difference between GFP fluorescence intensity of 3OC6HSL + aTc and that of 3OC12HSL + aTc. We referred this difference to our mathematical modeling.(Crosstalk Circumvention Assay)

Through this assay, we confirmed points below. ・lux/tet hybrid promoter is precisely repressed by TetR. This shows crosstalk circumvention. ・An affinity of 3OC6HSL-LuxR complex for lux/tet hybrid promoter is stronger than that of 3OC12HSL-LasR complex.
4. Mathematical modeling
Mathematical modeling of Ninja Circuit: Signal-dependent state change circuit with crosstalk circumvention
(detailed expression is here)
In order to clarify the parameter sensitivities and dynamic characteristics in the circumvention of crosstalk between intercellular molecules 3OC6HSL and 3OC12HSL, we modeled our circuit by using ODEs. We determined the transfer function of 3OC6HSL from the result of our assay of activation of our crosstalk circumvention system circuit (Fig. 2-1-19).
Furthermore, to know how much the circuit can circumvent crosstalk, we modeled our circuit in the presence or absence of the crosstalk circumvention system.
First, we considered the situation that E. ninja in the Attack state sees E. civilian . The following graph shows the switching from the Attack state to the Mimic state which is influenced by 3OC6HSL emitted by E. civilian. As you can see in the figure below (Fig. 2-1-20), when E. civilian comes at 300 min, the Attack state will switch to the Mimic state in around 100 min.
 Fig. 2-1-19. Activation of our crosstalk circumvention system circuit |
 Fig. 2-1-20. The change of each protein concentration in switching
|
During switching from the Attack state to the Mimic state, absence of TetR allows activation of lux/tet hybrid promoter. Protein TetR repressed by CI434, is indeed an important factor in the circuit. Under the Mimic state, TetR accumulates to plateau level. This presence of protein TetR is important to prevent crosstalk by 3OC12HSL-LasR complex.
Second, we considered the situation that when E. ninja is under the Attack state. When the Attack state turns into the situation of throwing shuriken, CI is expressed. Note that LacI expression form the lux/tet hybrid promoter is prohibited, due to the presence of TetR, even in the presence of 3OC12HSL-LasR complex which can bind to the hybrid promoter for its activation. After 3OC12HSL decomposition, the situation of throwing shuriken should go back to the Attack state (Fig. 2-1-21).
Interestingly, CI expression oscillates and converges by 3OC12HSL induction. This is not only because of the toggle switch, but also there is a repressilator by combination among TetR, LacI and CI434. Note that there is difference in CI concentration between the Attack state and the situation of throwing shuriken. This difference will be used for the decision to make E. ninja release shuriken or not.
We compared two circuits about the characteristics of changing from the LacI state to the CI state. The reason why we won’t compare them in the opposite way (changing from the CI state to the LacI state) is that crosstalk won’t occur in this direction. We analyzed the case that the initial state is with enough LacI, then it change to the state with enough CI at 300 min. by the increase of 3OC12HSL emitted from E. samurai. Fig. 2-1-22 shows the changing of LacI and CI. The solid line presents the case with crosstalk circumvention circuit, and the dotted line stands for the case without crosstalk circumvention circuit. When there is certain amount of 3OC12HSL production, LacI is produced in a certain amount in the toggle without crosstalk prevention circuit, and in contrary, the crosstalk would not be conspicuous. When E. samurai has gone at 1500 min., the concentration of LacI expressed by "Ninja circuit" is about 25 nM. In contrast, the concentration of LacI expressed by the original circuit is 725 nM. Regarding the convergence of LacI, we can conclude that LacI expressed by "Ninja circuit" will be converged faster than that of the original circuit does.

For confirming the efficiency of those two toggle switches, we set amount of LacI as the horizontal axis and CI as the vertical axis, and plotted out the changing. We can get the conclusion that the switching is much faster in the circuit with Ninja Circuit: Signal-dependent state change circuit with crosstalk circumvention.
5. Application
Our crosstalk circumvention system gives more flexibility to design genetic circuits because this system has a simple network topology composed of two repressor proteins, one repressor and one hybrid promoter. Along with the topology, you can just choose in any combination of sets of repressor protein and promoter. This system can be used for various genetic circuits to avoid other crosstalk.
6. References
- Timothy S. Gardner et al. (2000) Construction of a genetic toggle switch in Escherichia coli. Nature 403, 339-342
- Gray KM et al. (1994) Interchangeability and specificity of components from the quorum-sensing regulatory systems of Vibrio fischeri and Pseudomonas aeruginosa. Journal of bacteriology 176(10): 3076–3080.
- Hideki Kobayashi et al. (2004) Programmable cells: Interfacing natural and engineered gene networks. vol. 101 no. 22 8414–8419
