 "
"
Team:Peking/Project/BandpassFilter
From 2013.igem.org
| Line 262: | Line 262: | ||
#BPF_NewFig12{position:relative; left:250px; width:500px;} | #BPF_NewFig12{position:relative; left:250px; width:500px;} | ||
| - | #BPF_NewFig13{position:relative; left: | + | #BPF_NewFig13{position:relative; left:250px; width:500px;} |
#BPF_NewFig14{position:relative; left:250px; width:500px;} | #BPF_NewFig14{position:relative; left:250px; width:500px;} | ||
#BPF_NewFig15{position:relative; left:100px; width:800px;} | #BPF_NewFig15{position:relative; left:100px; width:800px;} | ||
Revision as of 20:29, 28 October 2013


Band-pass Filter
Introduction
Hitherto we have constructed a biosensor toolkit for aromatic compounds with wide sensing coverage and high orthogonality between different sensing modules. However, in order to cope with the need of in-field detection, we should further develop advanced equipment for our toolkit to implement fast, economical and convenient measurement of aromatic compounds in various environments.
Unfortunately, common reporting systems often failed to meet these requirements. This is because they often possess a Hill-function type dose-response curve. As can be observed from the dose-response curve of a typical Hill function (Fig. 1a), the linear range of a Hill function could be rather narrow, and the transition from low-output to high-output may be quite obscure to naked eyes. Thus appropriate equipment would be required to accurately measure output that follows Hill function type dose-response curve, making the measurement expensive and time consuming.

Figure 1. Dose-response curves and typical measurement results for a canonical reporting system a, and Band-pass Filter b. a, A general reporting systems typically possesses a Hill function type dose-response curve, and it's quite difficult to determine the absolute intensity of a particular signal among its gradually increasing outputs. b, dose-response curve of a Band-pass Filter possesses a single peak, and it's relatively easy to determine the position of the peak in its output series.
Although unaided eyes can barely determine the absolute intensity value of a particular signal among a series of signals with various intensities (Fig. 1a), humans are pretty competent at determining which signal is the strongest one, especially when there is a single peak among the signals (Fig. 1b). Thus it can be reasoned that if we are capable of transforming a series of signals with intensities changing monotonously into a series of signals with an unique intensity peak, reading and interpreting of the output signals will become much more intuitive and convenient. Fortunately, a Band-pass Filter is exactly the equipment that can turn a series of gradually increasing input signals into a series of output signals with a single peak.
It can be expected that when a Band-pass Filter is successfully constructed, we may serially dilute our sample into a concentration gradient and put our biosensor into the sample. The analyte concentration can be easily determined by a serial number of the test tube exhibiting highest output intensity (Fig. 2). We hoped that by implementing a Band-pass Filter circuit in our bacterial host cells, we might realize fast, economical and convenient detection of aromatic compounds in environment.

Figure 2. Graph illustration of proposed Band-pass Filter testing method. First a sample series need to be created by serially diluted the original sample. Then bacterial cells expressing Band-pass Filter circuits will be exposed to the sample series and the concentration of original sample will be determined based on serial number of the sample inducing highest output.
Concept of Band-Pass Filter
Band-pass Filter is a term used in electric engineering. It describes a device that passes signals with frequencies confined to a certain range and blocks signals with frequencies outside that range. The Band-pass Filter is constructed by combining a high-pass filter, which only pass signals with high frequencies, and a low-pass filter, which only pass signals with low frequencies (Fig. 3).

Figure 3. Sketch diagram of a typical Band-pass Filter in electric engineering. Vertical arrows show the input-output relationships of individual high-pass filters (left circle), and individual low-pass filters (right circle). The horizontal arrows show input-output relationship of a Band-pass Filter constructed by concatenating a high-pass filter and a low-pass filter. In an electric Band-pass Filter, the input signal is first processed by the high-pass filter to filter out low-frequency signals and then processed by the low-pass filter to filter out high-frequency signals, leaving only medium-frequency signals.
In analogy to an electric Band-pass Filter, a biological Band-pass Filter is a device that can be activated only by an input signal with medium intensity. Neither signal with low nor high intensity will generate an output signal (Fig. 4).
Quite similar to an electric Band-pass Filter, a biological Band-pass Filter can also be separated into two components, namely the two types of regulation the input node exerts on the output node in the network topology. In one way, the input node activates the output node through a positive feed-forward loop; in another way, the input node inhibits the output node through a negative feed-forward loop. Such a network topology, with two counteracting regulatory feed-forward loops connecting input node and output node, is called an incoherent feed-forward loop topology (Fig. 4a). The positive feed-forward loop will respond only to high intensity input signal (Fig. 4b),serving as a 'high-pass filter'. The negative feed-forward loop will respond only to the low intensity input signal (Fig. 4c), serving as the 'low-pass filter'. By fine-tuning transition points of the dose-response curves of the two counteracting feed-forward loops so that the transition point of the negative loop is higher than that of the positive loop, the biological Band-pass Filter, constructed by combining these two loops together, will respond only to a medium intensity input signal and generate an output peak at a specific concentration (Fig. 4d).

Figure 4. Sketch diagram of a possible topology (a) and functioning mechanism (b, c and d) of a biological Band-pass Filter. a, A network topology containing an Incoherent feed-forward loop, capable of generating a Band-pass Filter. The input node A directly represses output node C, creating a negative feed-forward loop, while indirectly activating output node C through repressing internode B which represses node C, creating a positive feed-forward loop. b, dose-response curve of positive feed-forward loop when characterized independently. The positive loop will respond only to high intensity input. c, Dose-response curve of negative feed-forward loop when characterized independently. The negative loop will respond only to low intensity input. d, The integrated dose-response curve of the incoherent feed-forward loop. High intensity input is filtered out by negative loop and low intensity input is filtered out by positive loop, only medium intensity input will induce a significant response.
Constructing Band-pass Filter
Having illustrated the basic principles of a Band-pass Filter, we set out to rationally design its genetic circuit.
First we selected three potential circuit networks (Fig. 5) with incoherent feed-forward loop as their core topology and then used Ordinary Differential Equations (ODEs) to analyze these circuit networks to identify the most robust circuit network. We chose to follow the four-node network because its performance remained more satisfactory than the others when the parameters varied randomly.

Figure 5. Graphs of three circuit networks we analyzed in our modeling. Each node represent a regulatory protein, either an activator or and repressor. All four networks possess incoherent feed-forward loops as core topology. Components of the activating half of an incoherent feed-forward loop are marked as green while components of repressing half are marked as black. a, b, Three-node networks taken into consideration. The principal difference between these two networks is the regulatory function of input node A. a, Three-node network where input node A functions as repressor. A directly repress output node C while indirectly activating it by inhibiting B, which represses C directly. b, Three-node network where input node A functions as activator. A directly repress output node C while indirectly activating it by activating B, which represses C directly. c, Four-node network taken into consideration. A indirectly activates output node D by activating C which activates D, while indirectly represses output by activating B that inhibits D.
Our next step is to select appropriate proteins to serve as individual nodes in the chosen circuit network. First we figured out the most crucial parameters in the ODE model through a parameter sensitivity analysis and determined the most desirable value for these parameters. Then we chose regulatory proteins whose kinetic parameter values are close to the desirable values, based on the reasoning that they would work much more efficiently than casually chosen ones.
Based on the analysis above, we selected phage transcription activator ϕR73δ as internode activator and phage transcription inhibitor cI as internode inhibitor while choosing NahR as the input sensor and sfGFP as reporter. The final construct is shown in Figure 6.
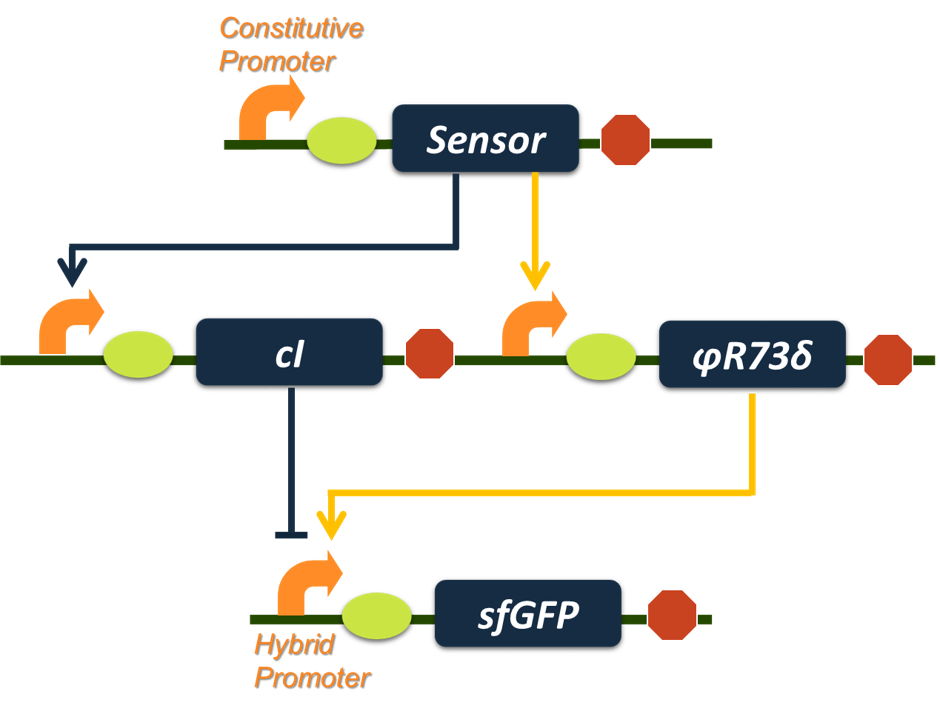
Figure 6.The final construct of our Band-pass Filter. The aromatic sensor (input node) will activate transcription of φR73δ and cI gene. The φR73δ will activate transcription of sfGFP reporter gene while cI represses transcription of the reporter gene, creating an incoherent loop. With proper parameter sets, such a genetic circuit will serve the function as a Band-pass Filter.
Details of constructing process can be viewed at Model page.
Building Our Hybrid Promoter
After determining the circuit network and protein candidates, we still need to address an important issue: we need to find a way to enable co-regulation of the reporter gene by two different transcription regulators. So we modified a bacteriophage ϕR73’s P2 promoter into a hybrid promoter that can be activated by the ϕR73δ activator and repressed by the repressor cI simultaneously and put reporter sfGFP under its regulation. We constructed the hybrid promoter by replacing the sequence between position -1 and -25 of P2 promoter with the cI binding site OR1 from Phage λ PR promoter. When ϕR73δ activator binds to its target sequence upstream of -35 element of the hybrid promoter, the transcription will start. The binding of cI dimers downstream of -35 element will block the binding of σ70 factors and thus repress the transcription even when ϕR73δ is bound. (Fig. 7).

Figure 7. Construction of Our Hybrid Promoter. Sequence information of phage φR73 P2 promoter (a), phage λ PR promoter (b) and our hybrid promoter (c) are shown. a, In the P2 promoter, φR73δ binds to a region between position -42 and -71 and activates transcription. b, In PR promoter, cI dimer binds to OR1 site (marked as blue) between position -9 and -25, blocking binding of σ70 factors and inhibiting transcription. cI binding region indicates the sequence we used to replace the corresponding region in P2 promoter.c, The hybrid promoter is constructed by replacing sequence between position -1 and -25 of φR73 P2 promoter with sequence at the same position in phage λ PR promoter that contains an OR1 site. The hybrid promoter is co-regulated by φR73δ and cI, with φR73δ activating and cI repressing. The repression of cI dominates over the activation of φR73δ, since the steric hindrance created by cI dimer prevents formation of transcription initiation complex even when RNA polymerases are recruited through the help of φR73δ.
In the Band-pass Filter circuit we constructed above (Fig. 6), the promoter will function in the following way as input intensity gradually increase: when the input intensity is weak, the concentration of ϕR73δ is too low to generate a strong output; when the input intensity is medium, despite a portion of promoters occupied by cI dimmers, the rest still can be activated by ϕR73δ and bring about a visible output; when the input intensity is strong, almost all of the promoters are blocked by cI dimers and the output is shut down. Hence only medium input signal can induce a significant output and the an single peak of output signal would be generated.
Characterizing Hybrid Promoter
As a key component of our Band-pass Filter circuit, the hybrid promoter must be carefully characterized in order to evaluate the feasibility of our Band-pass Filter circuit. To comprehensively characterize the dynamic performance of the hybrid promoter, we put two regulators of the hybrid promoter, ϕR73δ and cI, under the control of two different inducible promoters, Psal promoter and Ptac promoter. (Fig. 8). This enables us to manipulate separately the expression levels of two regulatory proteins through tuning Psal and Ptac promoter by adding different concentration combinations of inducers (salicylic acid for Psal promoter and IPTG for Ptac promoter).

Figure 8. Testing construct for hybrid promoter. φR73δ was put under the regulation of Psal promoter and cI was put under the control of Ptac promoter. Salicylic acid (SaA) will induce φR73δ expression and activate the hybrid promoter. Isopropyl β-D-1-thiogalactopyranoside (IPTG) will induce cI expression and repress the hybrid promoter. Expression level of the two regulatory proteins can be manipulated separately by adding different concentration combinations of SaA and IPTG.
To comprehensively characterized the hybrid promoter's transcription activity, we exposed the characterization circuit (Fig. 8) to a 8x8 two-dimensional induction assay established by combining 8 different concentrations of salicylic acid and 8 different concentrations of IPTG and measured the fluorescence intensity of sfGFP reporter using Flow Cytometry. (Fig. 9)

Figure 9. Characterization of hybrid promoter's dynamic performance. A two-dimensional inducer concentration assay was established by combining 8 different SaA concentrations (0, 0.1, 0.5, 1, 5, 10, 50 and 100µM) with 8 different IPTG concentrations (0, 1, 10, 50, 100, 150, 200 and 300µM). Bacteria cells expressing the testing construct were exposed to the assay and sfGFP fluorescence intensity was measured using Flow Cytometry. For a fixed IPTG concentration, fluorescence intensity gradually increased as SaA concentration increased. For a fixed SaA concentration , fluorescence intensity gradually decreased as IPTG concentration increased. These features indicated that the promoter functioned as expected.
The hybrid promoter worked as expected. For a fixed IPTG concentration, the sfGFP fluorescence gradually increased as the salicylic acid concentration increased, exhibiting a Hill-function type dose-response curve. For a fixed high salicylic acid concentration under which sfGFP expression is visibly induced, the fluorescence gradually decreased as the IPTG concentration increased, also exhibiting a Hill-function type dose-response curve. These data prove that the hybrid promoter can indeed be activated by ϕR73δ and repressed by cI, and the repressing effect of cI protein dominates over the activating effect of ϕR73δ protein, because transcription of the hybrid promoter can still be repressed to a very low level by cI even when ϕR73δ is expressed at a very high level. Simply characterizing the hybrid promoter won't satisfy us. We want to glean more information from this experiment in order to assess whether our Band-pass Filter design is really feasible or, in another word, whether the kinetic/dynamic parameter values of our genetic circuit actually fall within the range where a single output peak can be generated. However, there is an important feature in this testing construct that is radically different from our Band-pass Filter construct: the promoters driving the expression of ϕR73δ and cI are not the same, one is Psal, the other is Ptac. But this difference doesn't preclude the possibility of using data from this testing construct to give us insight on our original design. If the regulation mechanisms of the two promoters are close enough, we may reason that the Hill-functions describing the dynamic performance of the two promoters would also be similar (in the sense that their graphs can be overlapped by linearly stretching or compressing both axises). It is indeed the case. The Psal promoter is repressed by NahR tetramer through bending of DNA when salicylic acid is absent, and when salicylic acid is present, NahR will undergo a conformation change and transcription will start. (See Project, biosensors, NahR) Mechanism for Ptac promoter is rather similar: LacI inhibits transcription through tetramerization and DNA bending when lactose is absent and the inhibition is eliminated through conformational change. Following the reasoning above, we hypothesized that the negative feed-forward loop in the testing construct may actually represent a transformed version of the negative loop in the original Band-pass Filter construct. So we fit our model to the data from the testing construct in order to get real parameters for the Band-pass Filter circuit. (Fig. 10)

Figure 10. Model based data fitting for φR73δ activator (a) and cI repressor (b). a, Experimental points are sfGFP fluorescence intensities under different SaA concentrations without IPTG. The model based fitting curve provided parameter values for nA' and KAG. b, Experimental points are sfGFP fluorescence intensities under different IPTG concentrations along with 100µM SaA. Model based data fitting curve provided parameter values for nB', KBG and kAG•kBG. Fitting results: nA'=0.72705; KAG=62.99928; nB'=1.15498; KBG=11.53988; kAG•kBG=24671.78415. Definitions for the parameters can be viewed in equations written in Model page.
We substituted the parameters obtained from data fitting into the original Band-pass Filter to observe whether a peak is generated. (Fig. 11) Result showed that provided that our hypothesis is correct, our Band-pass Filter could indeed function as we expected.

Figure 11. Result of modeling based on parameters obtained from data fitting mentioned in Figure 10. Clearly a unique output peak is formed. This indicates that our band-pass filter circuit is feasible.
Based on the above model, we constructed the following genetic circuits. Because of the difference between Psal and Ptac promoters, precise data of the activation loop and repression loop, with their activator and inhibitor both controlled by Psal promoter, needed to be obtained.
Firstly, in order to measure the activation curve, the activator, ϕR73δ, was put under the regulation of Psal promoter placed on the low-copy-number plasmid pSB4K5 to create the regulator plasmid. (Fig. 12) A reporter plasmid, containing the hybrid promoter and downstream reporter gene sfGFP was also constructed using the high-copy-number plasmid pSB1C3 as backbone. The regulator plasmid was co-transformed with the reporter plasmid, and then the activation curve was measured. (Fig. 13)

Figure 12. Genetic circuit of the activation loop consisting of regulator plasmid (a) and reporter plasmid (b). The regulator plasmid contains the regulator NahR and the corresponding promoter Psal, which controls the activator ϕR73δ. The reporter plasmid contains the hybrid promoter Phyb and report gene sfGFP. The regulator plasmid and reporter plasmid use pSB4K5 and pSB1C3 as their backbone, respectively.

Figure 13. The induction ratio curve of the activation loop. X-axis is concentration of the ligand, 4-methylsalicylic acid, which activates the NahR. Y-axis is the induction ratio. It’s obviously that the induction ratio increases according to the increasing of the ligand.
Next, the inhibition effect of the repression loop also needed to be measured. The activator ϕR73δ was put under a constitutive promoter, Pc J23114 (Pc-114), to provide a high basal level of reporter expression. (Fig.14) Like ϕR73δ in the activation loop, cI was located downstream of the regulator NahR and the corresponding promoter Psal. The inhibitor and the activator were separated by a terminator, B0015, to prevent transcription leakage. The whole element was placed on the low-copy-number plasmid pSB4K5 as the regulator plasmid. The same reporter plasmid as in the activation loop is co-transformed with the regulator plasmid. The repression effect was measured. (Fig.15)

Figure 14. Genetic circuit of the repression loop consisting of regulator plasmid (a) and reporter plasmid (b). The regulator plasmid contains the regulator NahR and the corresponding promoter Psal, which controls the inhibitor cI. The activator ϕR73δ is expressed constitutively to provide a basal level of reporter gene. The reporter plasmid contains the hybrid promoter Phyb and report gene sfGFP. The regulator plasmid and reporter plasmid use pSB4K5 and pSB1C3 as their backbone, respectively.

Figure 15. The induction ratio curve of the activation loop. X-axis is concentration of the ligand, 4-methylsalicylic acid, which activates the NahR. Y-axis is the induction ratio. The repression effect has been displayed clearly. The induction ratio decreases according to increasing of ligand.
Reference:
[1] SOHKA, Takayuki, et al. An externally tunable bacterial band-pass filter.Proceedings of the National Academy of Sciences, 2009, 106.25: 10135-10140.
[2] MA, Wenzhe, et al. Defining network topologies that can achieve biochemical adaptation. Cell, 2009, 138.4: 760-773.
[3] BASU, Subhayu, et al. A synthetic multicellular system for programmed pattern formation. Nature, 2005, 434.7037: 1130-1134.
