|
|
From CRISPR to CRISPRi
CRISPR
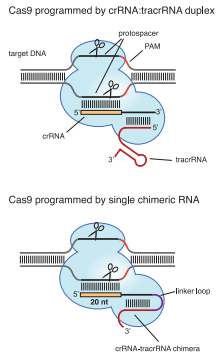 CRISPR Genome Editing (Jinek et al., 2012)
CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) and CRISPR-associated system (Cas) widely exist in Bacteria and Archea, endowing the cell with adaptive immunity to foreign DNAs. The immune process could be divided into three stages:
- Adaptive Stage. Fragments of foreign DNA (named protospacers) is incorporated into the proximal end of CRISPR.
- Expression Stage. Precursor CRISPR RNAs (pre-crRNAs) are transcribed from CRISPR. Pre-crRNAs are then cleaved to produce crRNAs.
- Interference Stage. crRNA complements with target DNA (protospacers), forming a complex that is recognized and bound by Cas nuclease (Csn). Csn excises the target, leaving the target vulnerable to successive degradation.
Three types of CRISPR system have already been revealed, of which Type II CRISPR system is the simplest – one sole protein, Cas9 (formerly named Csn1) is capable of all tasks once crRNA is complemented by trans-activating crRNA (tracrRNA). Due to this simplicity, Type II CRISPR/Cas can be ectopically expressed as a tool for sequence-specific genome editing (Jinek et al., 2012). And to further simplify the tool, Jinek et al. linked crRNA and tracr RNA together, successfully creating small guide RNA (sgRNA) as an equivalent (Jinek et al., 2012).
CRISPRi
With its endonuclease activity crippled, Cas9 can be used to hinder RNA polymerase and transcript elongation. Together with a proper sgRNA, Cas9 possessing such defect (named dCas9) can be used as an effective tool in expression interrogation – CRISPR interference, abbreviated as CRISPRi (Qi et al., 2013).
CRISPRi is promising tool for expression regulation, versatile and easy-to-use. Generally, a small shift in sgRNA would be enough for a new target.
Key Parts of CRISPRi
There are only 2 parts to be expressed -- dCas9 and sgRNA. Only 2!
dCas9
As the signature protein of the type II CRISPR/Cas, Cas9, does not show any detectable similarity to any proteins in Type I and Type III systems, in that it is sufficient both to generate crRNA and to cleave the target DNA. This large protein (about 1000 amino acids) contains two predicted nuclease domains -- the N-terminal RuvC-like nuclease (RNAse H fold) and the HNH (McrA-like) nuclease domain that is located in the middle of the protein (Makarova et al., 2011), each of which would cleave one DNA strand.
To acquire dCas9, one point mutation is conducted on each nuclease, respectively, namely, D10A and H841A (Qi et al., 2013).
sgRNA
If dCas9 acts as an executive, then sgRNA is the director of CRISPRi.
An sgRNA constitutes three parts:
A 20 nt complementary region for specific DNA binding.
a 42 nt hairpin for Cas9 binding
- A terminator derived from S. pyogenes, 42nt
Design Criteria (Qi et al., 2013)
- Binding specificity is determined by both sgRNA-DNA base pairing and a protospacer adjacent motif (PAM), NGG, upstream of target.
- Generally, the optimal length of the complementary region is 20nt.
- If a sgRNA targets coding sequence, it has to be complementary to the non-template (NT) strand. But if a sgRNA targets the promoter, either strand would be acceptable.
- If a sgRNA targets coding sequence, generally CRISPRi would work better if the complementary region is closer to the promoter.
- The first 7 nt might be a "seed region" for binding, since any single mutation of dramatically decreased repression.
References
JINEK, M., CHYLINSKI, K., FONFARA, I., HAUER, M., DOUDNA, J. A. & CHARPENTIER, E. 2012. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science, 337, 816-821.
QI, LEI S., LARSON, MATTHEW H., GILBERT, LUKE A., DOUDNA, JENNIFER A., WEISSMAN, JONATHAN S., ARKIN, ADAM P. & LIM, WENDELL A. 2013. Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression. Cell, 152, 1173-1183.
|
 "
"


