 "
"
Team:Marburg/Project:lightcontrol
From 2013.igem.org
m |
|||
| Line 4: | Line 4: | ||
{{:Team:Marburg/Template:ContentStartNav}}The registry of standardized biological parts consists of plenty of well characterized promoters. Many of these promoters enable the production of proteins for example antibodies after induction with a specific substance. However, using an external inducer leads to several drawbacks: The inducer must be removed from the medium firstly to inactivate the promoter and accordingly stop the expression and secondly to simplify the purification of the desired protein. | {{:Team:Marburg/Template:ContentStartNav}}The registry of standardized biological parts consists of plenty of well characterized promoters. Many of these promoters enable the production of proteins for example antibodies after induction with a specific substance. However, using an external inducer leads to several drawbacks: The inducer must be removed from the medium firstly to inactivate the promoter and accordingly stop the expression and secondly to simplify the purification of the desired protein. | ||
| - | Because plants and algae use sunlight as their primary energy source, they had to develop promoters, which respond to light. We therefore challenged the idea whether these light-inducible promoters would be suitable for regulating expression of target genes (Figure). The light-inducible promoter | + | Because plants and algae use sunlight as their primary energy source, they had to develop promoters, which respond to light. We therefore challenged the idea whether these light-inducible promoters would be suitable for regulating expression of target genes (Figure). The light-inducible promoter fcpB is used for the constitutive expression of proteins in genetically modified ''Phaeodactylum tricornutum'' algae. Unfortunately, this promoter is not well characterized until now. The fcpB-promoter is known to control the expression of fucoxanthin binding proteins. These proteins play an important role in the absorption of photosynthetic active sunlight. To protect the light harvesting complex (LHC) from dangerous high-energy sunlight these promoters are already induced by low levels of sunlight (Veith and Büchel, 2007, BBA). Therefore, we expected a promoter induction under blue, red and white light conditions and never under green light and in darkness due to the fact that photosynthesis occurs only when red or blue light is present and fucoxanthin binding proteins are necessary for light harvesting and photoprotection. That is the reason why we decided to study the promoter strength by radiating the cells with different transmission wavelengths (i.e. green (572 nm), blue (467 nm), red (672 nm), white light and darkness). Hence we used the light-inducible promoter for the expression of the reporter eGFP, which is advantageous because the expression level can be determined relatively easy. |
A ''P. tricornutum'' culture was grown for six days under the specific light conditions described above. Afterwards, cultures were kept in darkness for two days to minimize the amount of existing GFP in the cells (<html><a href="https://2013.igem.org/Team:Marburg/Notebook:Ptricornutum">see the protocol</a></html>). The optical density was measured and GFP was quantified by Western blot analysis. | A ''P. tricornutum'' culture was grown for six days under the specific light conditions described above. Afterwards, cultures were kept in darkness for two days to minimize the amount of existing GFP in the cells (<html><a href="https://2013.igem.org/Team:Marburg/Notebook:Ptricornutum">see the protocol</a></html>). The optical density was measured and GFP was quantified by Western blot analysis. | ||
Revision as of 17:46, 28 October 2013

Characterizing a new light-inducible promoter 
Because plants and algae use sunlight as their primary energy source, they had to develop promoters, which respond to light. We therefore challenged the idea whether these light-inducible promoters would be suitable for regulating expression of target genes (Figure). The light-inducible promoter fcpB is used for the constitutive expression of proteins in genetically modified Phaeodactylum tricornutum algae. Unfortunately, this promoter is not well characterized until now. The fcpB-promoter is known to control the expression of fucoxanthin binding proteins. These proteins play an important role in the absorption of photosynthetic active sunlight. To protect the light harvesting complex (LHC) from dangerous high-energy sunlight these promoters are already induced by low levels of sunlight (Veith and Büchel, 2007, BBA). Therefore, we expected a promoter induction under blue, red and white light conditions and never under green light and in darkness due to the fact that photosynthesis occurs only when red or blue light is present and fucoxanthin binding proteins are necessary for light harvesting and photoprotection. That is the reason why we decided to study the promoter strength by radiating the cells with different transmission wavelengths (i.e. green (572 nm), blue (467 nm), red (672 nm), white light and darkness). Hence we used the light-inducible promoter for the expression of the reporter eGFP, which is advantageous because the expression level can be determined relatively easy.
A P. tricornutum culture was grown for six days under the specific light conditions described above. Afterwards, cultures were kept in darkness for two days to minimize the amount of existing GFP in the cells (see the protocol). The optical density was measured and GFP was quantified by Western blot analysis. While determining the optical density, we found P. tricornutum growth under all light conditions except for darkness. Leblanc and coworkers showed that cryptochrome-, rhodopsin- and phytochrome-like receptors are present in marine diatoms indicating the ability to receive green light and to use green light for photosynthesis (Leblanc et al., 1999, Plant Mol Biol). Still, it was shown by Veith and coworkers that a fucoxanthin chlorophyll protein (FCP) complex, under the control of our light inducible promoter leads to a shift of the absorbance spectrum of PSI into the green spectrum after binding to fucoxanthin (Veith et al., 2007, BBA).
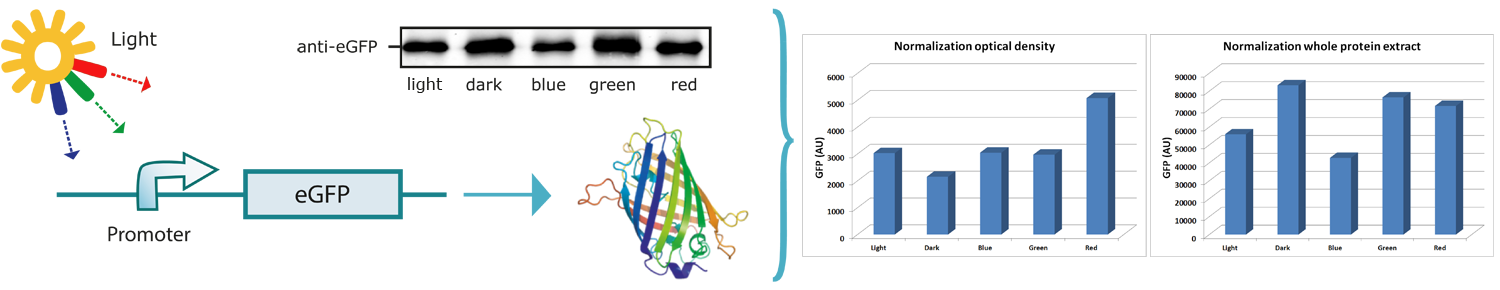
For the Western blot analysis, proteins were firstly extracted from the cells. As the standard method for cell disruption in P. tricornutum seemed too laborious and error-prone due to many intermediate steps, we decided to optimize the cell disruption in PHAECTORY. The methods to be tested were the disruption using glass beads, metal beads and ultrasound. As opposed to metal beads and ultrasound, glass beads did not lead to sufficient cell disruption. The first two methods yielded equal amounts of proteins. However the metal bead method seems to be preferable due to the fact that the ultrasonic method requires special equipment for the whole execution and the effects of the electromagnetic waves on the protein structure are not predictable. To minimize the protein decomposition by proteases the disruption was directly performed in SDS sample buffer containing 2-mercaptoethanol and immediately applied on a SDS gel (see protocol). The eGFP in the whole protein extract was detected using antibodies against the codon-optimized eGFP from Arabidopsis thaliana.
For the evaluation of the Western blot both, normalization against optical density (OD) and whole cell protein were done. In the case of the optical density normalization, it is clearly evident that light of any wavelength leads to the activation of the light-inducible promoter and thus increases the amount of eGFP in PHAECTORY. There is almost no visible difference in the amount of eGFP in the cells irradiated by blue, green and white light. In contrast, red light leads to more than 1.5 times the amount of eGFP. Hence, red light is the most efficient way to activate the light-inducible promoter fcpB in spite of the selective light absorption of longer wavelengths in water.
In the case of normalizing against whole cell protein, it can be observed that the highest amount of eGFP occurred in the cells exposed to darkness followed by the cells grown under green and red light conditions. This result does not agree with the one obtained by normalizing against the OD. Normalization against total cell’s protein amount might be hampered by the fact the amount of protein in the cell is not equal for the different light conditions tested. We therefore conclude that normalization against whole protein amount might not be appropriate. It could well be that in darkness the cells produce proteins, which are absent when grown under light or vice versa. However, further experiments have to be performed to understand these data. Taken together, we found that our promoter is light-inducible and can be activated by blue, green and red light whereof red light yields the best results.

